in
ГАМЕТЫ (греч. gamete — супруга), зрелые половые клетки, способные к оплодотворению.
При слиянии мужской и женской гамет образуется зигота, дающая начало новому организму. У мужчин постоянно в течение всей жизни, начиная с пубертатного возраста, образуется огромное число гамет (сперматозоидов), несущих примерно поровну Х- и Y-хромосом (сперматогенез). У женщин все половые клетки закладываются ещё внутриутробно, а в дальнейшем по достижении половой зрелости ежемесячно созревает одна яйцеклетка (овогенез). В связи с особенностями созревания гамет у женщин любое мутагенное воздействие на организм (начиная со стадии внутриутробного развития) может быть причиной генетических аномалий у её потомства. У мужчин, напротив, поколения спермиев довольно быстро сменяют друг друга, поэтому, если мужчина какое-то время находился в контакте с мутагенным фактором (например, служил на атомной подводной лодке), то спермии, образующиеся уже через 1-1,5 года после прекращения действия вредоносного фактора, не несут мутаций, связанных с радиацией.
Начало онтогенеза связывают с моментом образования зиготы, процессы же, предшествующие образованию зародыша — возникновение первично-половых клеток в организме, размножение и дифференцировка половых клеток — гаметогенез, а также оплодотворение,— объединяют понятием проэмбриональное развитие (прогенез).
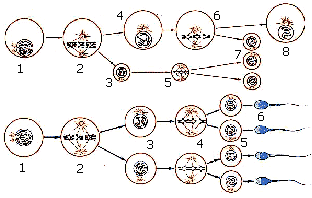
Схемa образования гамет (половых клеток) человека. Развитие яйцеклеток (вверху) и сперматозоидов (внизу):
Развитие яйцеклетки: 1 — в ядре ооцита первого порядка пары хромосом конъюгируют по всей своей длине; 2 — хромосомы выстраиваются в экваториальной плоскости и делятся мейотически; в результате образуется ооцит второго порядка (4) и первое полярное тельце (3), содержащие по половинному числу хромосом; 5 — хромосомы первого полярного тельца располагаются в экваториальной плоскости, готовясь к простому делению; 6 — в ооците второго порядка хромосомы располагаются в экваториальной плоскости, готовясь к простому (немейотическому) делению; в конечном счете образуется три полярных тельца (7) и зрелая яйцеклетка (8).
Развитие сперматозоидов: 1 — в ядре сперматоцита первого порядка происходит конъюгация хромосом по всей их длине; 2 — конъюгировавшие хромосомы выстраиваются в экваториальной плоскости и клетка делится мейотически с образованием двух новых клеток — сперматоцитов второго порядка, каждый из которых содержит половинное число хромосом; 3 — в ядре сперматоцита второго порядка хромосомы вновь конъюгируют по всей своей длине; 4 — хромосомы выстраиваются в одной плоскости для простого (немейотического) деления; 5 — из сперматоцита второго порядка образуются четыре предшественника сперматозоидов, или сперматиды; 6 — зрелые сперматозоиды.
При сравнении процессов сперматогенеза и оогенеза необходимо обратить внимание, во-первых, на то, что размножение сперматогониев пpoиcxoдит дольше и интенсивнее, чем размножение оогониев; во-вторых, рост ооцитов у многих животных идет несоизмеримо длиннее, чем рост сперматоцитов. Каждый сперматоцит дает четыре сперматиды, формирующиеся в четыре зрелых сперматозоида, в то время как каждый ооцит дает только одно зрелое яйцо.
Основные отличия половых клеток от соматических:
1. Сперматозоиды и яйцеклетки имеют не диплоидный набор хромосом, как это свойственно соматическим клеткам, а гаплоидный, т. е. в два раза уменьшенное число хромосом. Так, соматические клетки пчелы имеют 32 хромосомы, а сформированные половые клетки — 16. Соматические клетки человека имеют 46 хромосом, а сперматозоиды и яйца — 23.
2. У половых клеток резко измененное по сравнению с соматическими ядро-плазменное соотношение. Это очевидно на примере яиц птиц. Собственно яйцом (яйцеклеткой птицы) считается только «желток». Объем желтка яиц птиц в миллионы раз превышает объем клетки, исходной в его развитии. Объем же ядра столь резко не изменяется. И если из массы цитоплазмы яйца птицы исключить вещества, которые как бы включены в нее «про запас», для развития зародыша (желточные включения), и говорить о «чистой цитоплазме» (что не совсем правильно), все равно очевидно, что масса яйца сильно возрастает в объеме. То же наблюдается у яиц любого размера у всех животных, будь то большие яйца головоногих моллюсков, имеющих размеры до 10-15 мм в диаметре, 1,5-2-миллиметровые яйца гидры и лягушки или яйца млекопитающих, зародыши которых уже на ранних стадиях развития получают все необходимое от материнского организма. Например, у яйца женщины (100-300 мкм в диаметре) имеется также резко измененное соотношение ядра и цитоплазмы. У пиявки Nephelis ядерно-плазменное отношение в яйцевых клетках равно 0,15/428, а в соматических оно составляет 0,3/3 и менее. Значит, ядерно-плазменное отношение яйцеклетки по сравнению с телесными клетками в этом случае изменяется приблизительно в 300 раз. Впоследствии при развитии зародыша ядерно-плазменное отношение делящихся клеток восстанавливается до свойственного соматическим клеткам. Это происходит у разных животных в неодинаковое времся, но большей частью к 5-6-му или 7-му делению яйца.
Не менее «уродливые» изменения в соотношении цитоплазмы и ядра происходят при образовании мужских половых клеток. Как правило, сперматозоиды имеют очень мелкие размеры: у крокодилов, например, эти клетки около 20 мкм, у человека — 50-70 мкм . Об их ничтожных размерах можно судить, если представить, что сперматозоиды, оказавшиеся исходными в развитии всех современных людей (три миллиарда), по объему равны горошине. Имеются, впрочем, исключения: сперматозоиды одной из амфибий (Discoglossus pictus) — около 2 мм длиной. Объем ядер сперматозоидов несколько меньший, чем у соматических клеток, а «чистой цитоплазмы» у них очень мало. Грубо говоря, сперматозоид —.это ядро клетки плюс различные структуры, обеспечивающие его встречу с яйцом. Можно легко создать представление о соотношении ядра и цитоплазмы, если учесть, что головка сперматозоида почти целиком состоит из ядра.
3. Нормальные проявления жизни клетки, и прежде всего ее метаболизм, могут осуществляться лишь при тесной взаимозависимости ядра и цитоплазмы. Если мужские и женские половые клетки оказываются крайне измененными по сравнению с жизнедеятельными телесными клетками, то следует думать, что обмен веществ у них необычный. Высказано предположение, что яйцевая клетка в отношении обмена веществ находится в состоянии депрессии, которое можно уподобить анабиозу (спячке). Диссимиляторные и ассимиляторные процессы минимальны.
У сперматозоидов столь малое количество цитоплазмы и питательных веществ, что если бы им и был свойствен нормальный метаболизм, то он не мог бы продолжаться долго. В половых железах или в половых протоках самца сперматозоиды находятся в неподвижном и анабиотическом состоянии. Оказавшись вне мужской половой системы, они, как правило, живут очень короткое время. Так, сперматозоиды форели погибают в воде уже через 30 с, а у колюшки — через полчаса. В половых путях самок длительность жизни сперматозоидов у разных животных неодинакова. В яйцеводах кур сперматозоиды петуха живут 30-40 дней, у кролика 8-12 ч, в матке и яйцеводах женщины — 5-8 дней (а вне организма, в семенной жидкости — 2-3 ч). Имеются исключения. В семяприемнике самок пчел сперматозоиды сохраняют свою жизнеспособность 2-2,5 года, но они находятся в неактивном состоянии, с едва «теплющимся» метаболизмом. У летучих мышей спаривание происходит в начале осени. Оплодотворения, однако, не происходит, так как животные вскоре впадают в спячку, а с этим связано сильное понижение температуры тела и другие неблагоприятные для развития зародыша явления. Сперматозоиды сохраняются в течение всего зимнего периода в половых путях самки и весной, с возобновлением нормальной жизни, оплодотворяют яйцо.
Имеют место и паузы в развитии зародышей. Так, спаривание у косули Capreolus vulgaris происходит в августе, незамедлительно начинается развитие зародыша, но через несколько дней (на стадии бластоцисты — бластодермического пузырька) развитие останавливается до ноября, после чего возобновляется и идет беспрерывно до конца. Подобное явление наблюдается и у некоторых других животных (например, у соболя) и называется диапаузой. Несомненно, что большую роль при этом играют температурные условия. Например, при понижении температуры тела за 6-7 ч до 20°С у самок белых крыс на 4-й день беременности, когда зародыш находится на стадии бластодермического пузырька и еще не имплантируется в стенку матки, охлаждение в этот период вызывает состояние, подобное диапаузе, наблюдаемой у некоторых животных в естественных условиях. Возникает таким образом значительная задержка в имплантации.
4. Долгое время среди большей части биологов было распространено мнение о том, что половые клетки в отличие от соматических — это малодифференцированные «тотипотентные» клетки и что по мере развития организмов новые и новые клеточные поколения становятся все более и более дифференцированными. Затем стали считать, что это утвреждение неправильное и что яйца и спермии — высокодифференцированные клетки, обладающие многими специальными, выработавшимися в процессе эволюции приспособлениями для выполнения специфических функций: встречи мужских и женских половых клеток клеток — осеменения, защиты зародыша от инфекций и вредных абиотических факторов (оболочки яйцевых клеток). Яйцевые клетки каждого вида животных характеризуются особой структурой («проморфологией»), от которой зависят особенности развития организма; яйцевые клетки содержат те или иные вещества, требующиеся для метаболизма развивающегося эмбриона и т. д. Ядра яиц и сперматозоидов в генетическом смысле обеспечивают развитие признаков, свойственных матери и отцу.
Нормальные, жизнеспособные телесные клетки, содержа диплоидное число хромосом, при определенных экспериментальных или природных условиях могут быть аналогами половых клеток и развиваться в целый организм. Так происходит при бесполом размножении, естественной и экспериментальной полиэмбрионии, соматическом эмбриогенезе. Например, клетки листьев бегонии Begonia rex являются интегральной частью тканей, а при особых условиях они могут развиваться в новый организм с его воспроизводительными органами; при этом развившийся организм окажется с теми видовыми и индивидуальными наследственными свойствами, какие характеризуют и исходное растение.
5. Измененный по сравнению с соматическими клетками обмен веществ, наличие специальных структур, мешающих нормальному протеканию жизненного цикла клетки (ее онтогении), — все это приводит к тому, что сперматозоиды оказываются неспособными нормально, как это свойственно большинству соматических жизнедеятельных клеток, развиваться и доходить в своем развитии до итогового этапа онтогении клетки — митоза. Делящихся сперматозоидов ни у одного вида животных никтоиз ученых пока не наблюдал. Яйцевые клетки также не могут делиться без влияния особых внешних факторов, а именно: если не состоится оплодотворение или если яйцо не будет активировано к развитию партеногенетическими агентами (температура, механическое легкое повреждение оболочек (прокол), излучение и т.д.).
Зародышевый путь половых клеток
А. Вейсман в 1885-1896 гг. создал теорию «непрерывности зародышевой плазмы». Что существуют две плазмы: соматическая (сома), участвующая в развитии тела организма, и зародышевая (половая плазма), передающаяся непрерывно от половых клеток одного поколения к половым клеткам каждого нового поколения. Носителями жизни являются биофоры — группы молекул органических веществ. Они находятся в ядре клеток и через поры ядра могут переместиться в цитоплазму. Ядру принадлежит активная роль, а цитоплазма, ее поведение и дифференциация, структура и отправления зависят от детерминантов (определителей) — групп биофо. В зародышевой плазме находится полный набор детерминантов; их столько, сколько типов однородных клеток у взрослого организма. Детерминанты, как и биофоры, растут и размножаются (делятся), так что для любого числа однородных клеток определенной ткани достаточно одного исходного детерминанта. Все детерминанты, определяющие развитие всех признаков организма, сгруппированы в ядре в иды. Во время митоза иды слагаются в иданты, соответствующие структурам хромосом. Таким образом наступающие в развитии организма различия между клетками происходят путем сортировки наследственных единиц (детерминантов). В ходе эмбрионального развития в связи с делением клеток детерминанты распределяются по различным клеткам, происходит постепенная растрата их, обеднение ими у новых и новых поколений специализирующихся клеток. Только половые клетки имеют полный набор детерминантов, а потому они и способны развиваться в целый организм. На рубеже ХХ столетия перед Вейсманом вставали трудные вопросы. Соматические клетки по мере развития организма все более обедняются потенциями, растрачивают свои детерминанты. Каким же образом из обедненных телесных клеток возникают в яичниках и семенниках половые клетки с их полным набором детерминантов? В 1899 г. появилось исследование Бовери, которое, казалось, разрешало это тяжелое для теории Вейсмана противоречие. Бовери обнаружил, что уже первые два бластомера яйца лошадиной аскариды Ascaris megalocephala отличаются друг от друга. При втором дроблении в одном из бластомеров S1, исходном для эктодермы, происходит диминуция (уменьшение) хроматина благодаря отбрасыванию дистальных концов хромосом в цитоплазму; в другом бластомере Р1 - родоначальнике будущих внутренних, органов и половых клеток — диминуции хроматина нет. И при последующих дроблениях бластомеры Р1, Р2, Р3 и Р4 оказываются без диминуции, и именно эти клетки являются исходными для всех будущих сперматозоидов или яиц. Выходит, что уже на стадии 16 бластомеров имеется половой зачаток — гонобласт. А те бластомеры, которые делятся с диминуцией хроматина, являются поколениями соматических клеток. Это было триумфальным цитогенетическим подтверждением теории Вейсмана. Была обоснована теория особого зародышевого пути (Keimbahn) (линия половых клеток), разделявшаяся долгое время большинством эмбриологов. Было также доказано, что повреждение области половой плазмы (скопления ядер недимицированных клеток, плазмы линии половых клеток) (например, лучами Рентгена) ведет к развитию стерильных особей, что диминуция хроматина происходит под цитоплазматическим контролем и что половая плазма содержит фактор, защищающий хромосомы от диминуции.
Одни исследователи придерживаются представления о непрерывности в ряду поколений линии половых клеток, считая их развитие особым процессом, не имеющим подобия с другими явлениями дифференциации. Эти исследователи утверждают о существовании фундаментальных различий между соматическими и половыми клетками, состоящих в сохранении последними (как и их предшественниками в онтогенезе) качества тотипотентности — не дифференциации, а сохранения первоначального генома клетки без какой-либо существенной активации его частей. Возникновение половых клеток они связывают с теми клетками зародыша или организма, в которых в результате ооплазматической сегрегации окажется сосредоточенной зародышевая плазма, детерминирующая линию половых клеток и их предшественников в онтогенезе. Функция зародышевой плазмы предположительно состоит в предохранении генома клеток и их предшественников от соматизации, т. е. в обеспечении сохранения первичными половыми клетками их исходных потенций.
Другие исследователи считают, что формирование половых клеток тождественно процессам дифференцировки клеток в любых иных направлениях, а все особенности половых клеток связаны со спецификой их дифференцировки, обусловленной репродуктивными функциями: они отрицают непрерывность линии половых клеток в ряду поколений, но не отрицают реального факта раннего обособления их в онтогенезе многих животных, служащих единственным источником для образования половых клеток организма. Эти ученые утверждают, что под тотипотентность клеток обозначает скрытую, но сохранившуюся клетками способность дать при развитии целый организм. Такими потенциями при половом размножении Metazoa обладает зигота, а у животных с регулятивным типом дробления — бластомеры в ходе нескольких первых делений дробления. В эмбриогенезе по мере развития потенции всех клеток, не исключая и половые, все более суживаются (хотя утрата ими тотипотентности во многих случаях сопровождается сохранением эквипотенциальности ядер). При данном понимании тотипотентности ни первичные половые клетки, ни половые клетки всех этапов дифференцировки (ни гонии, ни гаметоциты, ни, тем более, зрелые половые клетки — яйца и сперматозоиды) не являются тотипотентными. С известной натяжкой можно говорить о тотипотентности зрелых яиц у некоторых животных в смысле их способности к партеногенезу; при этом следует помнить, что партеногенез при половом размножении — вторичное явление. Тотипотентность — это качество, воссоздаваемое каждый раз с появлением зиготы и утрачиваемое клетками зародыша, в том числе и половыми, в ходе их специализации. Половые клетки — это продукт организма, одноклеточного или многоклеточного, а их детерминация и дифференцировка всегда происходит под интегративным воздействием организма как целого.
При описании процессов эмбрионального развития, говорят дальше исследователи второго вида, можно говорить о линии половых клеток, но не надо вкладывать в это выражение метафизическое содержание, противопоставляя «бессмертную половую плазму» «смертной соме». У некоторых животных половой зачаток действительно образуется очень рано, но столь же рано обособляются и зачатки других органов и систем. На той же стадии развития аскариды, а именно на стадии 16 бластомеров, на которой обособляется половой зачаток, не менее отчетливо и обособление зачатка энтодермы (средней кишки). Таким образом, нет оснований придавать какое-то особое значение понятию «зародышевый путь» в отличие от «пути на эктодерму», от «пути на стомодеум» или от «пути на глаз».
Столь ранних случаев обособления полового зачатка, какое наблюдается у аскариды, очень немного {Sagitta, циклоп, некоторые насекомые). У большинства животных (у всех позвоночных) половой зачаток развивается сравнительно поздно. Вопросы происхождения половых клеток у животных связаны с конкретными, неодинаковыми для разных животных вопросами развития половых желез — семенников и яичников. Зачаток половых желез у животных возникает по-разному. Во многих случаях он образуется вместе с перитонеальными стенками целомическид мешков. У животных с ранним обособлением первичных половых клеток (оно происходит еще до возникновения мезодермы) перитонеальные клетки, покрывая слоем половые клетки, также принимают участие в развитии половой железы, являясь, в частности, источником образования фолликулярных клеток.
У губок нет непрерывности зародышевой плазмы: первичные половые клетки могут развиваться у них из амебоцитов, хоаноцитов или из археоцитов (из амебоцитов с крупным ядром). В отношении кишечнополостных также нет оснований говорить о «линии половых клеток». У гидр и других кишечнополостных нет постоянных семенников и яичников. Их половые клетки могут возникать из интерстициальных клеток, а также из зимогенных клеток гастродермиса. У полипа Clava squamata все половые клетки концентрируются в гонофорах. Если отрезать зону гонофора «с гроздьями» половых клеток под гипостомом, то при регенерации нового гипостома со щупальцами восстанавливается и весь половой аппарат за счет присутствующих в теле полипа интерстициальных клеток. Женские половые клетки у актинии Bunodactis stella образуются заново в каждый период половой активности непосредственно из эпителиальных клеток гастродермиса (септ). В оогенезе отсутствует стадия оогониальных делений. Зародыши и молодые животные не содержат клеток, которые можно было бы считать половыми. Созревание половых клеток происходит асинхронно; в одном мезентерии можно встретить ооциты, находящиеся на разных стадиях развития. У некоторых червей возможна регенерация гонад из участков тела, в которых отсутствуют половые элементы. Это наблюдается у планарий, а среди полихет — у серпулид; наблюдается это и у многих олигохет. У этих животных половые клетки обособляются на ранних стадиях эмбрионального развития, но соматические клетки способны восстанавливать половые элементы, которые утрачиваются в результате ампутации. У мшанок «обособление половых клеток» происходит на поздних стадиях развития. При бесполом размножении и регенерации из соматических клеток у них возникают половые клетки.
Членистоногие дают многочисленные и яркие примеры раннего обособления линии половых клеток. В поверхностном слое цитоплазмы (периплазме), в области заднего полюса имеется морфологически отличимый участок — «оосомы». При делении ядер, перемещении их в периплазму и формировании бластодермы особая судьба выпадает на долю тех ядер, которые окажутся в оосоме. Окружающая их цитоплазма разрастается, затем отделяется от остальной части яйца, образуя полярные клетки. У Drosophila melanogaster образование полярных клеток происходит на этапе развития зародыша, когда он состоит уже из 256 клеток и более. В цитоплазме заднего полюса яйца имеются какие-то условия, способствующие формированию гоноцитов (т. е. первичных половых клеток).
Пока не установлены надежные признаки первичных половых клеток у рыб, их происхождение остается неясным. Большинство авторов в настоящее время считают, что в течение всей жизни рыбы в половой железе сохраняются потомки первичных половых клеток, являющихся основным источником новых генераций как оогоний, так и сперматогоний.
Невозможно сомневаться в наличии линии половых клеток у всех позвоночных, однако споры по этому вопросу не прекращаются, к тому же эмбриология не располагает достаточными фактическими данными, например о рыбах.
Многие эмбриологи придерживаются «дуалистических» взглядов на развитие половых желез у позвоночных. Они считают, что соматические ткани яичника и семенника всегда мезодермального происхождения, а первичные половые клетки являются дериватами различных зародышевых листков и впоследствии мигрируют к зачатку половой железы (часто благодаря амебоидным движениям).
У амфибий первичные половые клетки, окруженные мезенхимой, могут быть обнаружены в микроскопических участках над формирующейся брыжейкой, откуда они перемещаются в мезодер-мальный эпителий, участвующий в развитии половой железы. У птиц и рептилий первичные половые клетки были найдены на ранних стадиях развития, перед началом сегментации мезодермы во внезародышевой желточной энтодерме. Клетки эпителиального типа обрастают здесь нераздробившуюся массу желтка. Они крупнее, чем обычные мезодермальные, содержат большое, более или менее вакуолизированное ядро, их протоплазма отличается от таковой окружающих клеток своим отношением к красителям, форма их близка к сферической. Сам по себе факт присутствия первичных половых клеток во внезародышевой желточной энтодерме еще не доказывает, что именно они или их потомки окажутся позднее составной частью функционирующей половой железы.
Считается, однако, доказанным, что «первичные половые клетки» мигрируют в мезенхиме и по кровеносным сосудам в область целомической мезодермы, где происходит закладка гонад. Располагаются ли «первичные половые клетки» в пределах зачаткового эпителия или во всей толще гонады? Гибнут ли вскоре все эти клетки, сыграв свою роль при формировании половой железы, или часть их становится родоначальником будущих половых клеток? Эти вопросы в достаточной степени еще не выяснены. Наиболее точными данными экспериментальная эмбриология располагает в отношении птиц и амфибий, и нет сомнений в том, что без мигрирующих в зачаток гонады «первичных половых клеток» яйца или спермин не формируются.
Несмотря на обилие исследований о развитии яичников и семенников у млекопитающих, вопрос о происхождении половых клеток у них далеко еще не ясен.
Согласно данным одних авторов, с момента прекращения митотического размножения первичных половых клеток у плода новообразования яйцевых клеток не происходит. Другие исследователи утверждают, что оогенез продолжается в течение всего репродуктивного периода жизни самки. Подсчитано, что число половых клеток у двухмесячного плода человека около 600 000. К пятому месяцу развития — 6 800 000. Позднее происходит массовая дегенерация клеток, так что ко времени рождения остается около 1 млн. нормальных ооцитов. Из этого количества примерно 300 000 сохраняется к семилетнему возрасту ребенка.
Многие ученые уверены в том, что первичные половые клетки полностью или частично дегенерируют и происходит формирование половых клеток из клеток герминативного эпителия. Описано новообразование яйцевых клеток из герминативного эпителия в разные периоды жизни (после рождения) у мышей, крыс, морских свинок, кошки, собаки, обезьяны и человека. Некоторые исследователи уверены в том, будто все клетки герминативного эпителия являются потенциальными яйцеклетками.
Однако, по данным многих авторов, после закладки половых желез ооциты не могут образовываться из клеток герминативного эпителия и абсолютно все ооциты происходят в результате размножения первичных половых клеток. Оогенез у млекопитающих, согласно мнению этих авторов, заканчивается на поздних стадиях развития зародыша или вскоре после рождения.
Исследования на позвоночных, зародышах человека позволили с полной уверенностью говорить о внегонадном возникновении первичных половых клеток у высших позвоночных, о миграции их в закладку гонады. Вследствие этого в отношении позвоночных надо отбросить понятие «зачатковый эпителий» и отказаться от мысли, будто есть эпителий — источник образования половых клеток. Имеет место раннее «обособление» линии половых клеток. Можно говорить о специальном половом зачатке — гонобласте. Существуют многочисленные данные о миграции первичных половых клеток, об активном движении гоноцитов через массу мезенхимных клеток (движение амебоидного характера), а также достоверные данные о переносе половых клеток с кровотоком. Местом первичной локализации гоноцитов эмбриона человека служит краниальная зона зародышевого щитка. Первичные половые клетки на разных стадиях развития человека оказываются в разных частях зародыша— они попадают в мезодерму первичной полоски, оказываются в составе кровяных островков, желточных сосудах, впоследствии в выстилке задней кишки, эндотелиальной закладке сердца и других образованиях.
В гонадах позвоночных гонии располагаются скоплениями — «гнездами». Позже вокруг каждой половой клетки за счет соматических элементов гонады формируется фолликул (от лат. folliculus — мешочек). Стенка фолликула состоит из фолликулярного эпителия, прилегающего к поверхности половой клетки и соединительно-тканной теки, в которой разветвляются кровеносные капилляры, питающие заключенную в фолликуле половую клетку. В оогенезе фолликул формируется по завершении гониальных делений и, как правило, содержит одну половую клетку — ооцит I порядка. В сперматогенезе формирование фолликула происходит задолго до завершения гониальных делений, продолжающихся внутри сформированного фолликула, благодаря чему семенной фолликул содержит большое количество мужских половых клеток. Кроме того, группы половых клеток внутри семенного фолликула могут быть заключены в сперматоцисты; стенка каждой сперматоцисты образована одной или несколькими фолликулярными клетками. Гомологом семенных фолликулов низших позвоночных у Amniota являются семенные канальцы. Находясь в контакте с половыми клетками, фолликулярный эпителий выполняет роль посредника в их взаимоотношениях с родительским организмом: регуляторных, трофических и иных. На определенных этапах гаметогенеза фолликулярный эпителий несет гормональную функцию; в случае гибели половых клеток осуществляет их фагоцитоз; продуцирует вторичные яйцевые оболочки.
Особенно велика роль фолликулярного эпителия в оогенезе в связи со значительным ростом ооцита и формированием яйцевых оболочек. В юных фолликулах фолликулярный эпителий состоит из одного слоя сильно уплощенных, функционально малоактивных клеток. На протяжении оогенеза фолликулярный эпителий позвоночных претерпевает значительные морфофизиологические превращения. Предложено различать четыре вида фолликулярных эгштелиев у позвоночных: однослойный статически плоский, в ходе оогенеза почти не изменяющийся (бесхвостые амфибии); трансформирующийся изоморфный, приобретающий в ходе оогенеза строение кубического, столбчатого или ложномногорядного (акулы, костистые рыбы, птицы); трансформирующийся анизоморфный, клетки которого в ходе оогенеза дифференцируются в двух направлениях — на собственно эпителиальные и железистые (скаты, чешуйчатые рептилии), и многослойный изоморфный — примером его служит фолликулярный эпителий плацентарных млекопитающих. Предполагается, что наиболее примитивным, исходным в эволюционных изменениях фолликулярного эпителия можно считать статически плоский, свойственный низшим вторичноротым. При овуляции стенка фолликула разрывается и яйцо освобождается в полость гонады либо в полость тела, откуда попадает в выводные пути половой системы.
Эволюция гаметогенеза
Исходным типом в эволюции гаметогенеза можно считать диффузный, солитарный (без участия вспомогательных клеток), распространенный среди губок, кишечнополостных, некоторых полихет, эхиурид. У некоторых многоклеточных диффузный гаметогенез осуществляется по алиментарному типу, т. е. с участием вспомогательных клеток, хотя и без гонад (некоторые губки, полихеты). У ряда низших первичноротых (некоторые кишечнополостные, приапулиды, некоторые черви и моллюски), хотя уже имеются простые мешковидные гонады, оогенез сохраняет черты солитарного типа, так как протекает в гонаде без участия вспомогательных клеток. Следующий этап в эволюции гонад многоклеточных связан с возникновением в них вспомогательных элементов гаметогенеза, образующихся в большинстве случаев за счет соматических клеток гонад (фолликулярные эпителии при фолликулярном типе алиментарного гаметогенеза) либо за счет некоторой части половых клеток («трофоциты» и др. — при нутриментарном типе алиментарного гаметогенеза).
В эволюции алиментарного оогенеза происходило усовершенствование трофических отношений женских половых клеток с организмом, обеспечивающих накопление яйцом резервных веществ. В эволюции алиментарного сперматогенеза происходила все большая изоляция развивающихся гоноцитов от внутренней среды организма. Особенно наглядно изолирующая роль фолликулярного эпителия в сперматогенезе может быть проиллюстрирована на примере сперматогенного эпителия млекопитающих. Клетки фолликулярного эпителия, выстилающие семенные канальцы, соединены межклеточным^ контактами такой структуры, что оказываются непроницаемыми для большинства веществ, находящихся в крови животного. Этот так называемый гематотестикулярный барьер имеется в семенниках млекопитающих, а также некоторых рыб и насекомых.
В целом эволюция алиментарного гаметогенеза осуществлялась по пути все большей специализации и расширения функции вспомогательных структур, достигших наибольшего развития у насекомых среди первичноротых и у позвоночных среди вторичноротых животных. О значении фолликулярного эпителия в гаметогенезе можно судить по перечислению его основных функций: это изолирующая, барьерная роль, функция транспорта, а иногда и синтеза трофических веществ длягоноцитов; гормонально-регуляторная, связанная с синтезом гормонов и гормоноподобных веществ, регулирующих гаметогенез; способность к фагоцитозу погибающих половых клеток; опорно-механическая и морфогенетическая функции; в овогенезе функция выработки вещества вторичной оболочки яйца и др.
Таким образом, характеризуя взаимоотношения половых клеток с организмом в филогенезе, можно отметить две противоречивые, но взаимодополняющие тенденции — все большую изоляцию половых клеток от внутренней среды организма и, с другой стороны, все большую зависимость развития половых клеток от вспомогательных элементов гаметогенеза.
Дополнительно:
Размножение
Эмбриональное развитие