|
|
|
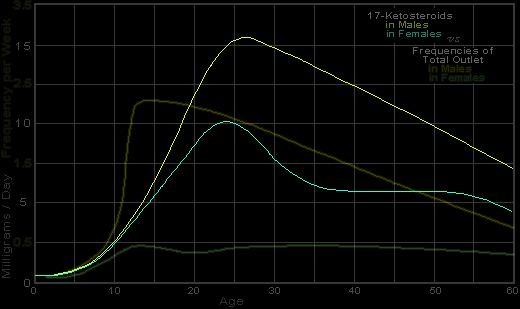
Figure 155f. Levels of 17-Ketosteroids in Human Female and Male |
|
Showing aging effect in male, and a prolonged plateau in the female, |
|
|
|
Figure 155f. Levels of 17-Ketosteroids in Human Female and Male |
|
Showing aging effect in male, and a prolonged plateau in the female, |