<<< >>>
{43}
Chapter 2
Humanistic Animals, Animalistic Humans
Titus and Ahab — male Gorillas — often courted and had sex with one another in the mountains of Rwanda, while Marchessa sought out her own sex during her pregnancy. In Florida, Bottlenose Dolphins Frank, Floyd, and Algie participated in homosexual activity with each other, as did Gabe and Moe-Miller, West Indian Manatees. Les and Sam (Siamangs) were doing the same in Milwaukee, while Kiku, a female Bonobo living in Congo (Zaire), had sex with her female "mentor" Halu more often than with anyone else in the new troop she joined. Cato and Mola (male Crested Black Macaques), Depp and Nice (male Rhesus Macaques), as well as Saruta and Oro (male Japanese Macaques) and Daddy and Jimmy (Crab-eating Macaques), also mounted one another. On the isle of Corsica, Le Baron and Le Valet (Asiatic Mouflons) were inseparable, as were Marian and her female Grizzly companion in the high mountains of Wyoming. Apolli and Arima — Long-eared Hedgehogs in Vienna — each refused to mate with males after they were separated from one another. In Austria, Greylag gander Pepino had a brief liaison with Florian but was later courted by Serge, while Max, Odysseus, and Kopfschlitz formed a threesome and went on to raise a family with Martina. A White-handed Gibbon named Floyd became sexually involved with George (his father) in Thailand, while Sibujong and Bobo, male Orang-utans, had sexual interactions with one another in Indonesia.1
As these examples show, zoologists sometimes bestow names upon the animals they study, lending an unintentionally — and eerily — human quality to their reports of homosexual activity. Although most scientists are careful to avoid anthropomorphizing their subjects, their use of human names such as these reminds us at once of each creature's individuality as well as the dangers of projecting human qualities onto animals. Such naming also demonstrates the nearly universal human preoccupation {44}
with seeking connections between ourselves and other species. Where animal behavior — especially sexual behavior — is concerned, it seems that comparisons will inevitably be made between animals and people (even by scientists).
There are a number of genuine connections and points of correspondence between animal and human homosexuality, as well as significant differences. There are also numerous pitfalls in attempting to extrapolate from animal to human behavior, or vice versa. This chapter explores a number of specific animal-human comparisons and the issues surrounding them. For example, we'll address claims that certain aspects of homosexuality are uniquely human, such as various types of sexual orientation, or the treatment of homosexual and transgendered individuals in the larger society. Also to be discussed are the special insights into human behavior offered by primate homosexuality (especially cultural behavior), and the rationale and motivations behind making cross-species comparisons in the first place (especially where the dubious concept of "naturalness" is concerned). Overall, a cautionary note must be sounded: while it is tempting to jump to broad conclusions about human homosexuality based on animal behavior (or vice versa), the full complexity and richness of homosexual expression in both animals and people must be considered. Only then will we begin to understand both the uniqueness and the commonalities of each.
From Pederasty to Butch-Femme: Uniquely Human?
One of the most significant results to emerge from the study of human homosexuality over the past few decades is the enormous variety of forms that this activity takes. From pederasty or "boy love" in ancient Greece, to ritualized homosexual initiation in New Guinea, to butch-femme lesbian relationships, to situational homosexuality in prisons, to contemporary North American gay couples — homosexuality has assumed many guises across history, cultures, and social situations. Thus, while homosexual desires and activities are probably ubiquitous, the specific forms that they assume are intimately shaped by particular sociohistorical contexts. Instead of talking about homosexuality, we should really speak in terms of homosexualities, plural, for there are many variations on the theme of same-sex relations.2
Animal homosexuality puts a new twist on this observation, since nearly every type of same-sex activity found among humans has its counterpart in the animal kingdom. Comparisons between animal and human homosexuality are inevitably muddled, however, by the lack of an adequate understanding and classification of different types of homosexuality. The confusion surrounding this subject is readily apparent: activities as different as pair-bonding between female Kangaroos and same-sex mounting in male Bighorn Sheep and Bottlenose Dolphins, for example, have all been compared to the sort of homosexual activity that occurs among human beings in prisons.3 The problem with analogies like this — which are inevitably imperfect and inaccurate — is that something like "prison homosexuality" is itself actually a conflation of many different behavioral variables and diverse patterns of same-sex activity, as are the "corresponding" animal behaviors.4 In addition to the actual form of the homosexual activity involved (pair-bonding, sexual behavior, {45}
etc.), many other factors must be considered, such as consensuality, age, gender presentation of partners, and so on. Thus, a particular example of homosexual activity — whether animal or human — is in reality a unique amalgamation or "blend" of multiple factors, any one of which may be shared with other forms of homosexual activity without necessarily conferring identity between the overall patterns they represent. Comparisons of homosexuality in animals and humans that fail to recognize such complexities are simply misleading.
It is helpful in this regard to think of homosexuality in terms of a number of independent axes, each of which is a continuum joining two "opposite" ends of a particular category (as suggested by researchers Stephen Donaldson and Wayne Dynes, who have developed a typology for human homosexuality based on this framework). 5 For example, one axis might represent the degree to which the homosexual interaction is gendered or role-based (ranging from the heavily role-oriented homosexuality of Native American two-spirits or Euro-American butch-femme lesbians, to the nongendered homosexuality of the South African San peoples or some gay couples in contemporary Euro-American culture). Another axis would represent the age relationship of the partners involved (ranging from no age difference to a clearly age-differentiated interaction); another represents sexual orientation of participants (homosexual - bisexual - heterosexual); another consensuality (forced or nonconsensual - freely chosen or consensual); another genetic relatedness of partners (incestuous - unrelated); another social status or position of same-sex activity (socially sanctioned - socially condemned); and so on.
The utility of such a system is that homosexuality in any given context (or species) can be seen as the intersection at various points on a number of such axes, thereby allowing comparisons to be made across multiple factors. In this chapter a number of these typological axes will be explored in greater detail to show that both animal and human homosexuality exhibit a comparable variability when examined from virtually every angle.6 Ultimately, we will see that the plurality of homosexualities in both animals and people suggests a blurring of the seemingly opposite categories of nature and culture, or biology and society. On the one hand, it is no longer possible to attribute the diversity of human (homo)sexual expression solely to the influence of culture or history, since such diversity may in fact be part of our biological endowment, an inherent capacity for "sexual plasticity" that is shared with many other species. On the other hand, it is equally meaningful to speak of the "culture" of homosexuality in animals, since the extent and range of variation that is found (between individuals or populations or species) exceeds that provided by genetic programming and begins to enter the realm of individual habits, learned behaviors, and even community-wide "traditions."
Comparisons between animals and people almost inevitably focus on behaviors that are supposed to be uniquely human. As biologist James Weinrich points out, nearly every behavior that was at one time believed to be practiced only by people has been found to have an analogue among animals — including homosexuality:
«There is a long and sordid history of statements of human uniqueness. Over the years, I have read that humans are the only creatures that laugh, that kill {46}
other members of their own species, that kill without need for food, that have continuous female receptivity, that lie, that exhibit female orgasm, or that kill their own young. Every one of these never-never-land statements is now known to be false. To this list must now be added the statement that humans are the only species that exhibit "true" homosexuality. Does anyone ever state that we alone exhibit true heterosexuality?»7
While many scientists now accept that animals engage in homosexuality, claims about human uniqueness continue to be made regarding the specifics of homosexual interactions: people, but not animals, engage in exclusive homosexuality, for example; people, but not animals, exhibit greater variety or "genuine" sexual motivation in their homosexuality; people, but not animals, react with hostility toward homosexuality and live in groups segregated by sexual orientation; and so on.
As we come to understand more and more about animal behavior, premature blanket statements like these have generally proven to be naive, if not incorrect — and this is especially true where homosexuality is concerned, since so much still remains to be learned regarding such activities in animals. In this section we'll address a number of these claims and explore some broader issues surrounding each (this theme will also be taken up in subsequent chapters with regard to the other typological "axes"). While there is some truth to these statements of human uniqueness, none is an absolute line of demarcation between human and nonhuman animals. As always, animal sexuality and social life are far more complex and nuanced than previously imagined: perhaps the only true difference in behavior between the species is that people, but not animals, are prone to make simplistic generalizations.
Exclusively Homosexual, Simultaneously Bisexual: Sexual Orientation
... preferential or obligatory adult homosexuality is not found naturally in any mammalian species other than Homo sapiens.
— W. J. GADPAILLE, 1980
Homosexual human couples who remain together throughout their adult life have few, if any, counterparts in wild mammals as far as is known at present.
— ANNE INNIS DAGG, 1984
Exclusive homosexual behavior appears to be absent among nonhuman primates ...
— PAUL L. VASEY, 19958
An oft-repeated claim about homosexuality is that exclusive, lifetime, or "preferential" homosexual activity is unique to human beings, or at least rare among animals (especially among primates and other mammals). This is really a question of sexual orientation — that is, to what extent do animals engage in sexual and {47}
related activities with members of the same sex without also engaging in such activities with members of the opposite sex? In fact, exclusive homosexuality of various types occurs in more than 60 species of nondomesticated mammals and birds, including at least 10 kinds of primates and more than 20 other species of mammals.9 In this section we'll consider these various forms of homosexual orientation and compare them to the wide variety of bisexualities that are also found throughout the animal world.
When discussing the question of exclusive homosexuality, several factors need to be distinguished: the length of time that exclusivity is maintained (short-term versus long-term, including lifetime), the social context and type of same-sex activity involved (pair-bonding versus promiscuity in nonbreeding animals, for example), the type of animal involved (e.g., mammal versus bird), and the degree of exclusivity (e.g., absolute absence of opposite-sex activity versus primary homosexual associations with occasional heterosexual ones, and vice versa). These factors combine in various ways and interact with each other to produce a number of different patterns. To begin with, we will consider long-term or extended exclusivity, since this pattern appears to be the most contested as to its existence among animals. Because species vary widely as to their life expectancy, onset of sexual maturity, and period of adulthood, it is difficult to come up with an absolute definition of long-term that has wide applicability. For the purposes of this discussion, though, we will somewhat arbitrarily consider homosexual activity that continues for less than two consecutive years (or breeding seasons) to be short-term, while anything continuing longer is considered extended or long-term, with the understanding that the latter category includes a wide spectrum of possibilities, anywhere from 3 years to a life span of over 40 years.
The only way to absolutely verify lifetime exclusive homosexuality is to track a large number of individuals from birth to death and record all the various homosexual or heterosexual involvements they have. Needless to say, this is a difficult task to accomplish (especially in the wild) and has been achieved for only a few species — indeed, in many cases the comparable evidence for lifetime exclusive
heterosexuality is not available either, for precisely the same reasons. Nevertheless, in at least three species of birds — Silver Gulls, Greylag Geese, and Humboldt Penguins — fairly extensive tracking regimes have been conducted, and individuals who form only homosexual pair-bonds throughout their entire lives have been documented. In some cases these are continuous pair-bonds that last upward of 15 years in Greylag Geese and 6 years in Humboldt Penguins (until the death of the individuals involved), while in other cases (e.g., Silver Gulls) individuals may also have several same-sex partnerships during their lives (either because of "divorce" or death of the partners).10
While absolute verification of lifetime homosexuality is not directly available for other species, extended periods of same-sex activity, perhaps even lifelong, are strongly suggested. In Galahs, Common Gulls, Black-headed Gulls, Great Cormorants, and Bicolored Antbirds, for example, specific homosexual partnerships have been documented as lasting for as long as six years (or individuals having several consecutive homosexual associations for that length of time); in most of these {48}
cases the absence of heterosexual activity for at least one partner has been documented or is highly likely. In many other bird species, same-sex partnerships that last anywhere from several years to life probably also occur: Black Swans, Ring-billed Gulls, Western Gulls, and Hooded Warblers, for instance. Although these durations have not been confirmed in specific individuals, homosexual pairs that continue for at least two years or birds who consistently form same-sex pairs for that time have been verified.11 In still other cases, long-term same-sex bonds undoubtedly occur because homosexual pairs in these species typically follow the pattern of heterosexual pairs, which are usually lifelong (or of many years duration): Black-winged Stilts, Herring Gulls, Kittiwakes, Blue Tits, and Red-backed Shrikes, among others. Finally, it must also be remembered that in many animals (e.g., Pied Kingfishers), same-sex (and opposite-sex) pair-bonds that last two to three years can still be lifelong, owing to the relatively short life span of the species.
In mammals, cases of long-term, exclusively homosexual pairing are indeed rare. One example is male Bottlenose Dolphins: the majority of males in some populations form lifelong homosexual pairs, specific examples of which have been verified as lasting for more than ten years and continuing until death. Although the sexual involvements (both same- and opposite-sex) of such individuals have not in all cases been exhaustively tracked, it is quite likely that at least some of these animals have little or no sexual contact with females (since breeding rates tend to be low in Bottlenose communities, with many individuals not participating in reproduction each year and, by extension, possibly throughout their lives).12 Absolute verification in this species, however, may not be forthcoming, since it is virtually impossible to continuously monitor the sexual behavior of all individuals within a given population of an oceangoing species. Bottlenose Dolphins are exceptional, however, in that the homosexual pattern in this species is distinct from the heterosexual one: opposite-sex pair-bonding does not occur among Bottlenose Dolphins. In most other species, homosexual and heterosexual activities tend to follow the same basic patterns, whether this means pair-bonding, polygamy, promiscuity, or some other arrangement.13 Lifetime homosexual couples are not prevalent among mammals, therefore, for the same reason that lifetime heterosexual couples are not: monogamous pair-bonding is simply not a common type of mating system in mammals (it is found in only about 5 percent of all mammalian species).14
Nevertheless, long periods of exclusive homosexuality among mammals have been documented for other social contexts besides pair-bonding. In many species, significant portions of the population do not engage in breeding or heterosexual pursuits for at least a part of their lives. Because some of these animals continue to engage in same-sex interactions, however, they are exclusively homosexual for at least that time, which can be considerable. Among Gorillas, for example, males often live in sex-segregated groups where homosexual activity takes place. The average length of stay in a male-only group is more than six years, although some males remain in such exclusively homosexual environments for much longer. One individual lived in an all-male group for ten years, staying until his death, and nearly a third of the males who joined the group over a thirteen-year study period were still with the group at the end of that time. Likewise, Hanuman Langur males may {49}
spend upward of five years in male-only bands in which homosexual activity takes place, and some individuals live their entire adult lives in such groups.15 In a number of hoofed mammals, a similar form of exclusivity based on sex segregation occurs: only a few individual males participate in heterosexual mating, while the remainder live in "bachelor herds" where homosexual activity often takes place.16 Among Mountain Zebras, for example, males stay an average of three years in such groups before joining breeding groups, and some remain their whole lives without ever mating heterosexually. Analogous patterns occur in a number of other species where only a relatively small percentage of males ever breed: antelopes and gazelles, including Blackbuck, Pronghorn, and Grant's and Thomson's Gazelles; Giraffe; Red Deer; Mountain Sheep; seals such as Northern Elephant Seals and Australian and New Zealand Sea Lions; and birds such as Ruffed Grouse, Long-tailed Hermit Hummingbirds, and Guianan Cock-of-the-Rock. In some hoofed mammals such as American Bison, a related age-based pattern is found. Males generally do not participate in heterosexuality until they are five to six years old; prior to that time, many engage in homosexual activities, entailing a period of exclusively same-sex activity of up to five years for some individuals.17
Other patterns of exclusivity occur as well. In Nilgiri Langurs and Hamadryas Baboons, for instance, generally only the highest-ranking male in a group mates with females; remaining males, if they engage in sexual activity at all, are sometimes involved only in homosexual pursuits. In Nilgiri Langurs, cases of nonbreeding males having only same-sex interactions for at least four years have been documented. In Ruffs, there are several different categories of males, many of whom rarely, if ever, mate heterosexually; some of these individuals participate in homosexual activities and may do so over an extended period, perhaps even for life. Finally, in some species same-sex activity may be exclusive because it is incestuous, involving a parent and its nonbreeding offspring. In male White-handed Gibbons, for instance, father-son sexual relations may continue for several years; the son is not involved in concurrent heterosexual activity, and sometimes even his father may have little or no opposite-sex mating during this time. Red Fox daughters can remain with their family group for many years — sometimes they never leave — during which time they may be involved in occasional same-sex mounting with their mothers (or each other) but no heterosexual activity.18
Thus, while in many species documentation of exclusive long-term homosexuality (or heterosexuality, for that matter) is not directly available, exclusivity can be inferred from the general patterns of social organization in the species. For example, a system that involves large numbers of nonbreeders (including individuals who never mate heterosexually during their entire lives), combined with homosexual activities among at least a portion of these nonbreeding animals (sometimes in sex-segregated groups), will invariably entail some individuals whose only sexual contacts are with animals of the same sex. For some animals this period of exclusive homosexuality lasts no more than a few years; for others, it may extend considerably longer, even for the duration of their lives.
Shorter periods of exclusive or "preferential" homosexuality also occur. Sexual "friendships" in Stumptail Macaques and Rhesus Macaques, for example, and {50}
homosexual consortships in Japanese Macaques, last anywhere from a few days to several months, during which time there are no heterosexual involvements. During the seasonal aggregations of male Walruses and Gray Seals, same-sex activity usually occurs to the exclusion of opposite-sex behavior. Female Marmots forgo breeding for a couple of years but may still have sexual contact with other females. Same-sex pair bonds in King Penguins and homosexual associations in female Orang-utans are also exclusive for their duration. Of course, many of these animals are actually bisexual because they also engage in heterosexual pursuits at other times during their lives, but while they are involved in same-sex activity, they do not simultaneously engage in opposite-sex behavior. Thus, when considering various forms of exclusive homosexuality it is also necessary to understand the different types of
nonexclusive homosexuality — that is, bisexuality.
The participation of an individual in both homosexual and heterosexual activities is widespread among animals: bisexuality occurs in more than half of the mammal and bird species in which same-sex activity is found. Nevertheless, there are many different forms and degrees of bisexuality, and these must be carefully distinguished when discussing sexual orientation in animals. A useful differentiation to start with is
sequential as opposed to simultaneous bisexuality, a distinction that hinges on the temporal or chronological separation between homosexual and heterosexual pursuits. In sequential or serial bisexuality, periods of exclusively same-sex activity alternate with periods of exclusively opposite-sex activity. In simultaneous bisexuality, homosexual and heterosexual activities co-occur or are interspersed within a relatively short period (say, within the same mating season). Thus, many of the "shorter" periods of exclusive homosexuality that we have been considering actually fall into a larger pattern of sequential bisexuality, which itself forms a continuum in which same-sex activity may occupy anywhere from several months to several decades of an animal's life. Moreover, the "sequentiality" of bisexual experience assumes many different forms: a seasonal pattern (for example, in Walruses, who engage in homosexuality primarily outside of the breeding season, or in {51}
Gray Whales, during migration and summering); an age-based pattern (e.g., in Bison or Giraffe, where same-sex activity is more characteristic of younger animals, or in which the earlier years of an animal's life are occupied largely with homosexual pursuits, to be followed by heterosexual activity in later years — or the reverse, as in some African Elephants); onetime "switches," in which individuals change over from heterosexual to homosexual activity at a specific point in time (e.g., Herring Gulls, Humboldt Penguins), or from homosexual to heterosexual (e.g., Great Cormorants); as well as less structured sequencing, in which several periods of same-and opposite-sex activity of varying lengths may alternate with each other (e.g., Gorillas, Silver Gulls, King Penguins, Bicolored Antbirds).19
Simultaneous bisexuality also assumes many guises. At one extreme, sexual activity with same-sex and opposite-sex partners takes place at
literally the same time: "pile-up" copulations, for example, in which a male mounts another male who is mounting a female (e.g., Wolves, Laughing Gulls, Little Blue Herons), or group sexual activity in which some or all participants are interacting with both males and females (e.g., Bonobos, West Indian Manatees, Common Murres, Sage Grouse). At the other extreme, individuals court or mate with both sexes separately, over short but relatively distinct spans of time, as in Crab-eating Macaques, Mountain Goats, Redshanks, and Anna's Hummingbirds. In between these extremes are other patterns, such as ongoing bisexual trios and quartets, in which both same-sex and opposite-sex partners are bonded to one another concurrently (e.g., Greylag Geese, Oystercatchers, Jackdaws). Another form of simultaneity involves an animal in a pair-bond with a member of the opposite sex who has occasional courtship and/or sexual encounters with a member of the same sex (or vice versa). For example, male Herring and Laughing Gulls, Herons, Swallows, and Common Murres who have female partners, and female Mallard Ducks who have male partners, sometimes mount birds of the same sex. Conversely, female Snow Geese, Western Gulls, and Caspian Terns and male Humboldt Penguins and Laughing Gulls who have same-sex partners sometimes mate with opposite-sex partners. Still another variation is found in Lesser Flamingos: males in homosexual pairs sometimes try to mate with females who are themselves in homosexual pairs. And in some animals such as Bottlenose Dolphins, Black-headed Gulls, and Galahs, the combinations are even more varied: different forms of sequential and simultaneous bisexuality, as well as exclusive homosexuality (and heterosexuality) are found in different individuals within the same species and may even combine in the same individual at different points in time.
Even within a given category of bisexuality — say, simultaneous bisexuality involving interspersed homosexual and heterosexual activity — each individual within a population generally exhibits a unique sexual orientation profile, consisting of his or her own particular combination of same- and opposite-sex activity. The concept of a scale or continuum as developed by Alfred Kinsey for describing human sexual orientation is useful here: within each species, individuals generally fall along a range from those exhibiting predominantly or exclusively heterosexual behavior, to those exhibiting a balance of both, to those exhibiting predominantly or exclusively homosexual behavior, and every variation in between.20 Species as a whole also differ as to where the majority of individuals fall along this continuum, {52}
and how many engage in more exclusive homosexuality or heterosexuality as opposed to more equal bisexuality. Thus, among Bonobos every female participates in both homosexual and heterosexual activity, but the proportion of same-sex behavior exhibited by each of the females in one particular troop varied between 33 percent and 88 percent (averaging 64 percent); in female Red Deer, from 0-100 percent (averaging 49 percent); among Bonnet Macaque males, between 12 percent and 59 percent (averaging 28 percent); in male Pig-tailed Macaques, from 6-22 percent (averaging 18 percent); and among Kob females, from 1-58 percent (averaging 11 percent).21 In other words, within an overall pattern of bisexuality, individual animals exhibit varying "degrees" of bisexuality — different "preferences," as it were, for homosexual as opposed to heterosexual activity.
These findings are particularly relevant since the concept of a scale or continuum of (homo)sexual behavior and orientation is yet another example of something still thought to be "uniquely human." The Bonobo data (as well as that for the other species) directly refute one primatologist's recent claim that "all wild primates we have seen within a particular species are equally homosexual ... . If you lined up ten female bonobos, it's not like one would be a 6 on the Kinsey scale and another a 2. They would all be the same number. It's only humans who adopt identities."
22 Of course, the Kinsey scale is specifically a measure of behaviors and
not identities (it was designed expressly to bypass the often problematic issue of people's "self-identification"), and certainly no animal studies purport to assess anything as subjective as sexual "identity." In its intended usage, though, the Kinsey scale (or a comparable measure of sexual gradations) in fact appears to be particularly apt for Bonobos. The figures cited above are based on the work of Dr. Gen'ichi Idani in Congo (Zaire), who studied a troop consisting of (coincidentally) exactly ten female Bonobos and tabulated all their homosexual genital rubbings versus heterosexual copulations over a three-month period. The percentages of homosexual activity in these individuals were 33, 36, 47, 68, 68, 70, 75, 75, 82, and 88 percent. Idani also tabulated the number of different male and female partners of each female (another possible measure of degree of bisexuality or behavioral "preference"). Again, the percentage of partners that were same-sex exhibits a range across all females: 36, 50, 50, 54, 67, 67, 67, 69, 71, and 80 percent. Clearly these individuals fall into a spectrum in terms of their sexual behavior and thus exhibit different degrees of bisexuality in terms of their sexual orientation (although none are in fact exclusively heterosexual or homosexual).23
"Preference" for same-sex activity is, admittedly, a rather elusive concept to measure when dealing with nonhumans (though not nearly as slippery as "identity"). Although we cannot access their internal motivations or "desires," animals do offer a number of other clues as to their individual "preferences" in addition to the proportion of their behaviors or partners that are same-sex. These include homosexual activity being performed in (spite of) the presence of members of the opposite sex, individuals actively competing for the attentions of same-sex partners (rather than "resorting" to such activity), advances of opposite-sex partners being ignored and/or refused, and "widowed" or "divorced" individuals continuing to pair with same-sex partners after the loss of a homosexual mate (even when {53}
opposite-sex partners are available). These types of behaviors have in fact been reported in more than 50 mammals and birds (see the profiles for some examples), indicating that for at least some individuals in these species, same-sex activity has "priority" over opposite-sex activity in some contexts. The converse is also true for species such as Canada Geese, Silver Gulls, Bicolored Antbirds, Jackdaws, and Galahs: in situations where opposite-sex partners are not available, only a fraction of the population engages in same-sex activity, indicating more of a heterosexual "preference" in the remainder of the population.24 Animals who do participate in same-sex activity in such a situational context could perhaps be said to exhibit a "latent" bisexuality; i.e., a predominantly heterosexual orientation with the potential to relate homosexually under certain circumstances. Another factor to be considered when evaluating individual "preferences" or degrees of bisexuality is the consensuality of the sexual interaction. Female Canada Geese and Silver Gulls in homosexual pairs, for example, may engage in occasional heterosexual copulations under duress; i.e., they are sometimes forcibly mated or raped by males. Likewise, heterosexually paired males in Common Murres, Laysan Albatrosses, Cliff Swallows, and several Gull species may be forcibly mounted by other males. Technically, all such individuals are "bisexual" because they engage in both homosexual and heterosexual activity, but the sort of bisexuality they exhibit is far different from that of a female Bonobo or a male Walrus, for instance, who willingly mates with animals of both sexes.
Broad patterns of sexual orientation across individuals show almost as much variation as that within individuals. In some species, the majority of animals are exclusively heterosexual, but a small proportion engage in bisexual activities (e.g., Mule Deer) or exclusively homosexual activities (e.g., male Ostriches). In others, the vast majority of individuals are bisexual and few if any are exclusively heterosexual or homosexual (e.g., Bonobos). Other species combine a pattern of nearly universal bisexuality with some exclusive homosexuality (e.g., male Mountain Sheep). In still other cases, the proportions are more equally distributed, but still vary considerably. In Silver Gulls, for instance, 10 percent of females are exclusively homosexual during their lives, 11 percent are bisexual, and 79 percent are heterosexual. Homosexual-bisexual-heterosexual splits for specific populations of other species include: 22-15-63 percent for Black-headed Gulls; 9-56-35 percent for Japanese Macaques; and 44-11-44 percent for Galahs.25
Thus, sexual orientation has multiple dimensions — social, behavioral, chronological, and individual — which must all be taken into account when assessing patterns of heterosexual and homosexual involvement. It is true that exclusive homosexuality in animals is less common than bisexuality — but it is not a uniquely human phenomenon, for it occurs in many more species than previously supposed. Moreover, because of the wide prevalence of bisexuality — both within and across species — exclusive
heterosexuality is also certainly less than ubiquitous. Animals, like people, have complex life histories that involve a wide spectrum of sexual orientations, with many different degrees of participation in both same- and opposite-sex activities. To the question "Do animals engage in bisexuality or exclusive homosexuality?" we must therefore answer "both and neither." There is no such {54}
thing as a single type of "bisexuality" nor a uniform pattern of "exclusive homosexuality." Multiple shades of sexual orientation are found throughout the animal world — sometimes coexisting in the same species or even the same individual — forming part of a much larger spectrum of sexual variance.
Nonchalant Onlookers and Gay Ghettos: Social and Spatial Responses
While homosexual behavior is widespread among our primate relatives, aggression specifically directed toward individuals that engage in it appears to be a uniquely human invention.
— PAUL L. VASEY26
An aspect of animal homosexuality that has received little attention in both popular and scientific discussions is the position or "status" of homosexual, bisexual, and transgendered individuals in the larger society. What kind of social response do they evoke from the animals around them? What is their spatial relation to the rest of the population — are they segregated, fully integrated, or somewhere in between? Primatologist Paul L. Vasey suggests that homosexual behavior in primates is characterized by a noticeable lack of hostility and segregation from the animals around them, and by and large this does appear to be true — not only for primates, but also for the vast majority of other species in which homosexual activity occurs. Almost without exception, animals with "different" sexualities and/or genders are completely integrated into the social fabric of the species, eliciting little of the attention, hostility, segregation, or secrecy that we are accustomed to associating with homosexuality in our society. Observer after scientific observer has commented on how homosexual behavior in animals is greeted with nonchalance from nearby animals. Individuals move effortlessly between their homosexual activities and other social interactions or behaviors without eliciting so much as a second glance from the animals around them.27
Where individuals engaging in homosexual activity do attract attention, it is usually out of simple curiosity (e.g., African Buffalo, Musk-oxen), or else because other animals want to participate.28 In a number of species such as Bonobos, Killer Whales, West Indian Manatees, Giraffes, Pronghorns, Common Murres, and Sage Grouse, homosexual interactions between two animals often develop into group sessions as more and more animals are drawn to the activity and join in. This is also true for heterosexual interactions in many of these species, and sometimes homosexual and heterosexual activity are part of the same group interaction. This illustrates an important point concerning the integration of homosexual activity within the larger social framework: when bisexuality is prevalent in a species, or when a large proportion of the population engages in homosexual activity (as is often the case), the distinction between "homosexual" and "heterosexual" animals melts away, as does the potential for aggressive responses based on those categories. An "observer" of homosexual activity could just as easily be a participant at some other time, and any separation between animals that engage in same-sex activity and those that don't becomes essentially arbitrary.
{55}
Even in species where homosexuality, bisexuality, or transgender are not widespread, animals that participate in same-sex behaviors (or transgendered individuals) are not generally treated to adverse reactions from the majority around them. Rather, homosexual activity is regarded as routinely as heterosexual activity is. In fact, in many species it is heterosexual, not homosexual, behaviors that draw a negative response. In numerous primates and other animals, for example, male-female copulations are regularly harassed and interrupted by surrounding animals. Same-sex activity in these species is either disregarded altogether (e.g., Stumptail Macaques), or else is subject to a much lower rate of harassment and interruption than opposite-sex matings (e.g., Hanuman Langurs, Japanese Macaques).29 Adult male Bonobos interfere with the heterosexual pursuits of younger males while ignoring (or even participating in) their homosexual activities, while heterosexual breeding pairs of Jackdaws, rather than same-sex pairs, are sometimes terrorized by nonbreeding heterosexual pairs (who may even kill their young). And in Guianan Cock-of-the-Rock, heterosexual courtship interactions are routinely interrupted and harassed by other males while homosexual activities are not. In fact, females defer to males engaging in same-sex courtship or copulation (by leaving or avoiding the display grounds while this activity is going on), and males may actually interrupt heterosexual interactions by initiating homosexual ones.30
Not only are homosexuality and transgender largely devoid of negative responses from other individuals, in some cases they actually appear to confer a positive status on the animals involved. In species that have a ranked form of social organization, for instance, homosexual activities are often found among the highest-ranking individuals (e.g., Gorillas, Bighorn Sheep, Takhi, Gray-capped Social Weavers). Likewise, transgendered animals sometimes have high status in a population (e.g., Savanna Baboons) or are more successful than other animals at obtaining sexual partners (e.g., Red Deer, Common Garter Snakes).31 While the benefits experienced by these individuals are not necessarily a direct result of their transgender or homosexuality, in a few cases individuals actually do appear to rise in status or obtain other positive results specifically because of their homosexual activities. Black Swans and Greylag Geese who form homosexual partnerships, for example, often become powerful, high-ranking forces in their flocks, in part because the combined strength of the paired males gives them an advantage that single males and heterosexual pairs do not have. In fact, Black Swan male pairs sometimes acquire the largest and most desirable territories in their domain, relegating other birds to a distinctly disadvantaged status.32
Same-sex couples in many animals routinely defend their home territories against intruders or assist their partners in conflicts with other individuals (as do heterosexual pairs).33 However, some homosexual and transgendered individuals in a number of species take this a step further, not merely defending themselves but actually going on the offensive. Gander or cob pairs often become so powerful that they are able to "terrorize" an entire flock, attacking individuals (as in Greylag Geese) or even forcing heterosexual pairs to give up their nests and eggs (as in Black Swans), which they take over to raise as their own. Male pairs of Flamingos have also been known to steal nests from other birds, while single males occasionally {56}
pursue and harass heterosexual pairs out of interest in the male (rather than the female) partner. Female pairs of Orange-fronted Parakeets often behave aggressively toward heterosexual pairs and may actively "dominate" them through attacks and threats, even successfully competing against them for possession of nesting sites. Laughing Gull homosexual pairs sometimes intrude on territories belonging to neighboring heterosexual pairs and harry the owners, as do pairs of male Rose-ringed Parakeets. A similar pattern has also been reported for Nilgiri Langurs, in which two males who live in the same troop and sometimes participate in same-sex mounting with each other (without necessarily being bonded to one another) may cooperate in attacking males from neighboring troops. Male Lions who are involved in homosexual courtship and sexual activities may attack other males who get too close to them, leading to intense fights in which the courting pair is often assisted by other group members who are not themselves directly involved in the homosexual activity. Homosexual consortships between female Rhesus Macaques can develop into powerful and highly aggressive alliances when the partners take the initiative in attacking other individuals and even driving them from the troop; female Japanese Macaques often vigorously compete with males (and other females) for access to female sexual partners. One female Common Chimpanzee that had sexual relationships with other females was consistently aggressive toward other individuals and came to be feared by Chimps of both sexes. Sometimes the aggression is directed at rival heterosexual partners: a female Livingstone's Fruit Bat who sexually pursued her own mother, for example, successfully fought off males that were also interested in mating with her mother. Finally, a transgendered Savanna (Chacma) Baboon was one of the strongest and highest-ranking members of her troop; described as exhibiting "courage and determination," she routinely interfered in heterosexual matings by threatening, "capturing," and then "carrying off" the male partner so she could mate with him.34
Ironically, then, some of the most aggressive interactions surrounding variant sexualities/genders in animals involve heterosexual individuals being attacked, harassed, or invaded by homosexual, bisexual, and transgendered individuals. Nevertheless, the converse situation is not unknown: there are a number of examples of homosexual animals being targeted by heterosexual ones. All such cases involve a male interfering with homosexual activity between two females, often in an attempt to gain sexual access to one of the females. Male Brown Capuchins, Rufous Rat Kangaroos, and Sage Grouse occasionally try to break up mating activity between females, while male Gorillas have been known to attack two females that are having sex together. A male Bonobo repeatedly tried to interfere in the sexual activity between females by screaming, jumping, and sometimes even hitting them; rather than preventing their sexual interactions, however, this simply caused the females to have sex with each other furtively until he gave up harassing them, after which they could do so openly. Male Canada Geese and Wapiti sometimes try to separate female pairs and mate with one member by driving the other away or isolating her from her companion (the females usually do manage to get back together), while female Japanese and Rhesus Macaques in homosexual consortships are occasionally threatened and charged by males. Jackdaw females who are bonded {57}
to each other as part of a bisexual trio may be hindered in their joint parenting efforts by their male partner, who sometimes prevents one of the females from having access to their nest. In some cases, this may lead to a loss of eggs or young.35 Notably, these reaction patterns are not typical for most of these species, since on other occasions animals usually have no adverse response to same-sex activity (e.g., in Bonobos, Gorillas, Rufous Rat Kangaroos, and Sage Grouse). And rarely do these attempts at interference (even when violent) force individuals to permanently cease homosexual activities: rather, they simply alter their patterns of relating or resume their activities once the interference has stopped.
In contrast, there is a consistent pattern among White-tailed Deer of highly aggressive attacks against the transgendered "velvet-horns" (individuals who combine both male and female characteristics). These animals are continually hounded by nontransgendered Deer of all ages and sexes, who drive them away and prevent them from approaching feeding grounds. Sometimes a "gang" of up to half a dozen bucks will attack a velvet-horn, charging, chasing, and severely wounding it with their antlers. Possibly as a result of this social ostracism, velvet-horns tend to associate only with other velvet-horns, forming their own groups and generally avoiding other Deer.36 Other than this example, though, it is rare to find animals with different sexualities or genders living separately because of persecution from members of their own species.
In many cases, animals that are involved in homosexual interactions do live in segregated groups, but their spatial and social separation from other individuals is based on factors other than their sexuality (since such groups typically also contain individuals who do not engage in homosexuality). Among such factors are age, sex, breeding status, social rank, activity patterns, and various combinations of these. For example, homosexual activity is characteristic of groups of younger, nonbreeding, and/or lower-ranking Northern Elephant Seals; of nonbreeding males in the sex-segregated "bachelor herds" characteristic of many hoofed and marine mammals; of groups of Cliff Swallows engaged in mud-gathering activities away from the nesting colonies; of older, solitary African Elephant males; of nonbreeding Pied Kingfishers who are not involved in helping heterosexual pairs; and of groups of male Gray Seals who gather together during the molting season. Physical disabilities can also isolate individuals into their own groups: in Greenshanks, for instance, flocks of one-legged birds have been observed socializing and migrating separately from other individuals. This is probably because they are unable to keep up with other birds rather than because of social ostracism, since two-legged birds are also sometimes found in such flocks.37 In contrast, although some Greenshanks participate in homosexual activity, no corresponding "flocks" of clearly homosexual or bisexual birds are known in this species.
Still other factors besides hostility from other animals may be involved in the occasional segregation of individuals that participate in homosexual activity. Among Ring-billed Gulls, for instance, female pairs are sometimes relegated to poorer-quality nesting sites or smaller territories, or they end up congregated together in the spaces between territories belonging to heterosexual pairs. Although this could be due to active hostility from neighboring birds, it is just as likely due to {58}
the fact that female pairs are generally not as aggressive as male-female pairs and consequently are unable to defend their nest sites from the encroachments that all pairs must endure in crowded colonies. In addition, heterosexual pairs of Ring-billed Gulls that are younger or less experienced also tend to end up in suboptimal locations, and in some colonies female pairs are fully integrated or randomly distributed rather than being peripheralized or clustered. This indicates that hostility toward female pairs is not ubiquitous and, if present at all, is not directed exclusively toward female pairs. In some species segregation is actively initiated by the individuals who are involved in homosexual activity. Female Japanese Macaques in homosexual consortships, for instance, isolate themselves physically and socially from other troop members, including their relatives, to spend time together. Likewise, Black Swan male pairs may end up physically separated from other individuals, but this is because their territories are the most expansive and also because they are aggressive toward other birds that approach them. Greylag gander pairs tend to occupy a peripheral position in their flocks, but it is unlikely that they have been "forced" to the edges, since male pairs are typically more domineering than any other birds in the flock. Some scientists have suggested that such homosexual pairs may actually be performing the role of "sentinels" or guards for the group as a whole, hence their position at the flock's "border." There is also evidence that homosexual Mallard Ducks prefer each other's company and tend to congregate together: when large numbers of male pairs were brought together in captivity, for instance, they tended to form their own flocks and socialize with each other rather than with heterosexual birds. The reason such flocks are not often seen in the wild, then, may simply be a matter of numbers. Because same-sex pairs tend to make up a minority of the population in this species, it is unlikely that enough homosexual individuals would ever be present together in a wild flock to form their own large groups.38
Nevertheless, the virtual absence of segregated subgroups of homosexual or bisexual individuals in the animal world is probably related at least in part to the general lack of overt hostility toward homosexuality among animals. Of course, {59}
multiple factors are undoubtedly involved, as is true for the formation of segregated groups of homosexuals among people. The emergence of "gay ghettos" or subcultures in some human societies is a complex process related to many things besides gaining refuge from persecution, such as the need to find and associate with one's own kind, the formation of a homosexual "identity," the development of economic independence, and so on. Nor are such groups merely a defensive response to a hostile society: as with many other minorities, such "ghettos" may begin as a necessary survival tactic but then develop into vital and enriching subcultures of their own. For animal societies we have already seen that many other factors — widespread bisexuality, for instance, or small numbers of animals participating in same-sex activities — can mitigate against the formation of separate groups. Conversely, segregated social units in which homosexual activity takes place often form for reasons that are (initially) unrelated to the sexuality of the animals involved. However, it is striking that both active hostility toward individuals involved in homosexuality and segregation of such individuals are rare occurrences in the animal kingdom. While neither of these social responses to homosexuality is uniquely human (as has been claimed), they are generally uncharacteristic of animal societies. Homosexuality, bisexuality, and transgender are usually as much a part of animal social life as heterosexuality, regardless of their prevalence or frequency of occurrence. In this respect, the vast majority of other creatures have an approach to sexual and gender variance that is decidedly
humane, rather than human — and they might even offer us models of how our societies could integrate differently oriented or ambiguously gendered individuals into the fabric of social life.
Sexual Virtuosos: Heterosexuality and
Homosexuality Compared
The complete pattern of human heterosexuality is not found in any other species (social-class differences in sexual behavior, pair-bonding, face-to-face copulation, hidden menstruallestrous cycles, oral and anal intercourse, etc.), although any single aspect of human heterosexuality can be found in some animal species. The same statement can be made about human homosexuality.
— JAMES D. WEINRICH39
It is ironic that this last assertion about how human beings are unique in their sexual behavior should have been made by the same scientist who commented on how rarely such statements of human uniqueness prove to be true. Indeed, now that more detailed and comprehensive information is available about animal homosexuality, it appears that at least three species rival, if not equal, human beings in the variability and "completeness" of their sexual expression: Bonobos, Orang-utans, and Bottlenose Dolphins. For each of the features mentioned above, an identical or equivalent aspect of behavior can be found, at least in a same-sex context. While none of these species has rigidly stratified "social classes," there are discernible differences in sexual behavior between animals of different ages and social statuses. Homosexual activity is often more frequent in younger, lower-ranking {60}
female Bonobos who have recently joined a new troop, for instance, while younger individuals are often "on the top" during female homosexual interactions and "on the bottom" during male homosexual interactions. There is also some evidence that sexual activity between females occurs more often when they belong to distant rank classes. Adolescent or younger-adult Orang-utans participate in same-sex activities more than older, higher-ranking individuals and also exhibit distinctive heterosexual patterns. Younger male Bottlenose Dolphins tend to form their own groups in which same-sex activity is more common than among pair-bonded individuals — while adult males in this species generally do form lifelong bonds with each other.
Although Bonobos do not have exclusive pair-bonding per se, females do form long-lasting bonds with each other that include sexual interactions; adolescent females also typically pair up with an older "mentor" female when they arrive in a new group and engage in sexual activity most frequently with her. Orang-utans often form pairlike consortships in heterosexual contexts, and similar sorts of associations also occur in homosexual contexts. Sexual interactions between female Bonobos usually occur in a face-to-face position, as do heterosexual (and some homosexual) interactions in Orang-utans, as well as most copulations in Bottlenose Dolphins (for the latter, a "belly-to-belly" position is perhaps a more apt characterization). "Hidden estrous cycles" refers to the fact that no overt physical changes signal the various phases of a woman's sexual cycle. A female Bonobo's sexual skin swells with her cycle, although it is present for the majority of the cycle and is not associated specifically with ovulation. Bottlenose Dolphins do not generally give any visual cues as to their sexual cycles or timing of ovulation, nor do female Orang-utans.40 In any case, all three species engage in sexual activity throughout the female's cycle the way human beings do, which is one important consequence of concealed sexual cycles in humans. Finally, Bonobos, Orang-utans, and Bottlenose Dolphins all engage in forms of anal and oral sex (the latter including fellatio in Bonobos and Orangs, cunnilingus in Orang-utans, and beak-genital propulsion in Dolphins).41
One area relating to sexual variability where animal and human homosexuality have been claimed to be comparable, rather than different, concerns the variety of sexual acts or positions used in homosexual as opposed to heterosexual contexts. Masters and Johnson found that gays and lesbians in long-term relationships often had better sexual technique and more variety in their sex lives than married heterosexuals. James Weinrich has suggested a parallel to this observation among animals, claiming that animals that engage in same-sex activity are, in a sense, sexual "virtuosos," employing a wider range of sexual behaviors, positions, or techniques than do their heterosexual counterparts.42 Although the validity of this claim with respect to human beings cannot be directly addressed here, its accuracy with respect to animals can be assessed — and it appears that in this case the situation is considerably more complex than previously supposed.
It is certainly true that homosexual repertoires have a wider range of sexual acts than heterosexual repertoires in a number of species. In Stumptail and Crab-eating Macaques, for instance, oral sex and mutual masturbation occur primarily, if not exclusively, between same-sex partners. Male Bonobos have a form of {61}
mutual genital rubbing known as penis fencing that is unique to same-sex interactions. Male West Indian Manatees employ a wider variety of positions and forms of genital stimulation during sex with each other than do opposite-sex partners. Various types of anal or rump stimulation (besides mounting or intercourse) occur in homosexual but not heterosexual contexts in several Macaque species, Siamangs, and Savanna Baboons. Only same-sex partners participate in reciprocal mounting in at least 15 species, including Bonnet Macaques, Mountain Zebras, Koalas, and Pukeko.43
This does not, however, appear to be part of an overall pattern, especially where sexual activities other than mounting or intercourse are concerned. For example, of the 36 species (exhibiting homosexual behavior) in which some form of oral sex is practiced, in only 10 (28 percent) of these is oral-genital stimulation limited to homosexual contexts, and in some cases (e.g., Rhesus Macaques, Caribou, Walruses, Lions) genital licking is a uniquely heterosexual act. Similarly, manual stimulation of the genitals or masturbation between partners is limited to same-sex interactions in 15 of the 27 species (55 percent) where this behavior occurs; in the remaining animals, both heterosexual and homosexual (or, in some cases, only heterosexual) partners are involved. Even anal or rump stimulation (besides intercourse) is found in heterosexual contexts in half of the species (6 of 12) that engage in such activities. Combining these observations, we find that a variety of sexual acts are part of both heterosexual and homosexual repertoires in the majority of cases, with behaviors unique to same-sex interactions occurring only in about 40 percent of the cases.
As a matter of fact, in most instances both heterosexual and homosexual acts are equally "uninspired," involving nothing more exotic than mounting behavior in the front-to-back position typical of mating in most animals. Even considering animals where other mounting positions are used, however, it is overly simplistic to claim that same-sex activity involves more "versatility." In many species a variety of positions are employed in
both heterosexual and homosexual situations. Furthermore, even though their frequency of use differs depending on the context, the major distinctions in mounting positions often lie along lines of gender rather than sexual orientation. The differentiating factor is not whether sexual activity involves partners of the same or opposite sex, but whether it involves males (in either heterosexual or homosexual contexts). For example, a face-to-face position is used for roughly 99 percent of sexual interactions between female Bonobos, but rarely in male-female interactions. However, a face-to-face position is almost equally rare in male homosexual interactions, occurring in only about 2 percent of activity between males. Thus, male homosexuality is more similar to heterosexuality than either is to female homosexuality in terms of the frequency of use of these two basic positions. A similar pattern occurs in Gorillas: although the face-to-face position is virtually absent in heterosexual encounters and much more common in homosexual ones, the two sexes have almost opposite preferences for this position. Almost three-quarters of female homosexual encounters involve the face-to-face position, while more than 80 percent of male homosexual mountings involve the front-to-back position (also preferred for heterosexual encounters). In Hanuman Langurs, {62}
male homosexuality is also more similar to heterosexuality than is female homosexuality in terms of the way that interactions are initiated: both males and females typically invite males to mount them by performing a special "head-shaking" display, which is much less characteristic of mounting between females.44
In Japanese Macaques, female homosexuality is more similar to heterosexuality than to male homosexuality in terms of the
variety of positions used. In contrast, male homosexuality is more similar to heterosexuality than female homosexuality in terms of the
frequency that various positions are used. In this species, fully seven different mounting positions can be identified, including four varieties of the front-to-back posture (with the mounting animal sitting, lying, or standing — with or without clasping its partner's legs — behind the mountee), two types of face-to-face positioning (sitting or lying down), and sideways mounts. All seven of these positions are found in both heterosexual and female homosexual encounters, while male homosexual mounting employs only five of the seven (sitting or lying on the partner in a front-to-back position are not used). However, sexual encounters between females differ from both heterosexual
and male homosexual mounts in using the face-to-face position more often, and in using the double-foot-clasp posture less than 20 percent of the time (compared to 75-85 percent of the time for sexual encounters where males are involved, either heterosexual or homosexual).45
Other patterns based on the intersection of gender and sexual orientation also occur. In Stumptail Macaques, for example, female homosexual encounters use three basic positions (standing or sitting front-to-back and sitting face-to-face), heterosexual activity uses two of these (standing or sitting front-to-back), while male homosexual activity uses only one of these (standing front-to-back). (Males do, however, employ a wider range of oral and manual forms of genital stimulation in their encounters with each other.) Copulations between female Flamingos generally resemble heterosexual matings more than they do mountings between male Flamingos, but same-sex copulations in birds of either sex differ from heterosexual activity in their lack of a particular "hooking" posture.46 Male White-handed Gibbons interact sexually with other males only in a face-to-face position and with females only in a front-to-back position — thus, homosexual and heterosexual interactions are equally "flexible" or "inflexible" in this species, but differ in which position is preferred in each context. Even reciprocal or reverse mounting — in which partners take turns mounting each other — is part of the heterosexual repertoire in more than three-quarters of the species that engage in this activity (in either same-or opposite-sex contexts); it is unique {63}
to heterosexual relations in many of these (including Western Gulls and Silvery Grebes) and present in many animals that do not engage in homosexual behavior at all.
In fact, it is sometimes the case that opposite-sex partners show
more variability or flexibility in their sexual activity. Heterosexual copulation in Botos occurs in three main positions (all belly-to-belly, either head-to-head, or head-to-tail, or at right angles) while homosexual copulation usually uses only one of these (head-to-head).
47 Both heterosexual and homosexual encounters in this species can involve two different forms of penetration (genital slit or blowhole), although same-sex activity also includes a third option, anal penetration. Among birds, the overwhelming majority of species mate in the standard position of one individual mounted on the other's back, in both heterosexual and homosexual contexts. The only examples of other positions being used with any regularity involve male-female mounts: a facing position (extremely unusual for birds) is used by stitchbirds, for instance, who mate with the female lying on her back and the male on top of her, and in purple-throated Carib hummingbirds, who mate belly-to-belly while perched on a branch. Copulation in red-capped plovers is achieved by the male first throwing himself on the ground on his back, then pulling the female on top of him in a facing position. Vasa parrots have an elaborate and (for birds) unusual form of genital contact in which the male inserts his genital protrusion (a bulbous swelling surrounding his genital orifice) inside the female's cloaca, which extends and envelops his organ while the two birds transfer from a regular mounting position to a side-by-side position (full penetration does not usually occur in bird matings). Vervain hummingbirds actually mate in midflight while traversing an 80-foot trajectory low above the ground. Finally, several species of woodpeckers are true heterosexual virtuosos: in an acrobatic sequence the male first performs a standard front-to-back mount and then drops to one side of the female, making genital contact with his tail underneath hers and sometimes ending up on his back or with his entire body in a perpendicular or even upside-down position.48
"Virtuosity" in other areas of behavior is not generally exclusive to homosexual encounters either. The vast majority of courtship interactions, for example, involve the same set of behaviors typical for the species regardless of whether they are being performed between partners of the same or the opposite sex. There are notable exceptions, of course: the courtship "games" of female Rhesus Macaques and solicitations of female Japanese Macaques; "necking" interactions between male Giraffes; pirouette dances in male Ostriches; the vocal duets of Greylag gander pairs; aspects of courtship feeding in Laughing Gulls, Antbirds, Superb Lyrebirds, and Orange-fronted Parakeets; alternative bower displays in Regent Bowerbirds; and unique vocalizations during homosexual but not heterosexual interactions in male Emus and Japanese Macaques. Occasionally courtship activities are also performed at different rates or with different intensities: in same-sex pairs of Black-winged Stilts and Black-headed Gulls, for instance, certain courtship behaviors occur more frequently in same-sex pairs, others more commonly in opposite-sex pairs. All of these represent behavioral innovations in same-sex contexts, but they are atypical. Usually both homosexual and heterosexual courtships draw upon the same repertoire {64}
of behaviors, and in many cases same-sex interactions actually involve only a subset of the full behavioral suite that is characteristic of the species.
Thus, while homosexuality among animals is sometimes characterized by innovative or exceptional behaviors not found in heterosexual interactions, the opposite situation is equally, if not more, prevalent. It seems, then, that neither virtuosity nor mundanity of sexual expression are exclusive to either homosexual or heterosexual contexts. This is really not surprising: as we have already seen, a hallmark of sexual (and related) behaviors in animals is the tremendous range of variation found between species as well as among different individuals. For just about any pattern or trend that can be discerned, one that is contradictory or equivocal can be found. It stands to reason, then, that something like "sexual technique" would exhibit a similar range of diversity. And although Masters and Johnson may have found a greater level of technical proficiency in sex among some homosexual couples, this is probably an overly simplistic generalization even among people. A wider study sample that includes extensive cross-cultural information, as well as closer attention to age, gender and class differences, social contexts, and other factors, would likely reveal that (once again) human beings are much more like other species in this regard.
Primate (Homo)Sexuality and the Origins of Culture
Homosexuality is part of our evolutionary heritage as primates: anyone looking at the prevalence and elaboration of homosexual behavior among our closest relatives in the animal kingdom will be led, eventually, to this conclusion. In fact, primatologist Paul L. Vasey traces the occurrence of homosexuality in primates back to at least the Oligocene epoch, 24-37 million years ago (based on its distribution and characteristics among contemporary primates).49 Some of the most organized and developed forms of homosexuality among animals can be found in the more than 30 species of monkeys and apes where this behavior occurs. Bonobos, for instance, engage in both male and female homosexual interactions with disarming frequency and enthusiasm, and they have also developed many unique forms of sexual expression, including a type of lesbian tribadism known as genito-genital rubbing. Similar elaborations of homosexual patterns are found among Stumptail Macaques, Gorillas, Hanuman Langurs, and many other monkey and ape species. In addition to highly developed systems of same-sex interaction and diverse sexual techniques, a number of other aspects of homosexual activity in primates are particularly salient. Among these are various forms of pair-bonding such as consortships, "favorite" partners, or sexual friendships; evidence for exclusive or preferential homosexual activity in some individuals (as discussed in the preceding section); female orgasm in monkeys and apes, in both homosexual and heterosexual contexts; female-centered or matrifocal societies, as well as male alliances and other groups of cooperating males in some species; and the wide range of nonreproductive heterosexual activities found in many primates.50
In addition to being part of our evolutionary heritage, homosexuality is also part of our cultural heritage as primates — for same-sex activity in monkeys and {65}
apes offers us some startling examples of cultural traditions among animals. Although "culture" is something that we typically associate with human beings, many animals innovate behaviors and then pass them on from generation to generation through learning. Zoologists speak of this as "cultural" behavior in animals — or, if the activity is less well developed, as "precultural" or "protocultural" behavior. Animal cultural traditions are widespread and often highly complex, occurring in many different kinds of species and involving behaviors as diverse as foraging and hunting techniques, communication patterns and song dialects, forms of social organization, response to predators, characteristics and locations of home sites and shelters, and migration patterns.51 Perhaps the most famous example of animal cultural behavior concerns food-gathering techniques in wild Japanese Macaques: in the mid-1950s, one female invented several ways of accessing novel food items (introduced by investigators), including sweet-potato washing, peanut digging, and "placer mining" for wheat. Within ten years 90 percent of all troop members had acquired these habits, which were being learned spontaneously by younger animals and passed on to subsequent generations.
"Culture" can also involve social behaviors: male caretaking of infants in Japanese Macaques, for example, is characteristic of only certain populations and appears to be a learned behavior, acquired by some individuals or troops and not others. Sexuality — including homosexual activity — can also bear the hallmarks of cultural activity. Scientists studying mounting behavior by females — once again in Japanese Macaques — suggest that whether and how females mount male or other female partners may represent a form of protocultural behavior. Certain mounting positions, for example, seem to become more "popular" in some troops over time, only to wane and be replaced by others. Likewise, masturbation among females appears to be learned through observation or other social channels. Although a
capacity for homosexual activity (along with reverse mounting and masturbation) is probably an innate characteristic of the species (as evidenced by at least some level of these activities in most populations), its occurrence between different troops and individuals is highly variable. Key aspects of such activities are apparently being learned and passed on through space and time. This indicates that "traditions" or patterns of sexual activity may be innovated and then transmitted via a web of social interactions, moving between and within population groups, geographic areas, and generations. Sexuality, including aspects of both same-sex and opposite-sex interactions, is also considered to exhibit aspects of cultural traditions in at least two other primates, Stumptail Macaques and Savanna Baboons.52
Not only is sexuality itself a form of cultural behavior, it can also impact and intersect with other sorts of cultural innovations in primates, often in surprising ways. In fact, nonreproductive sexual activities, including homosexual behaviors, may have contributed to the development of a number of significant cultural "milestones": hallmarks of evolutionary and cultural change that are considered to be defining characteristics of "humanness," yet which also exist in prototypical form in some of our primate relatives (and presumably also in our protohuman ancestors). In this section we'll briefly consider the role that sexuality might have played in the development of primate communication systems and the origin of language, {66}
in the manufacture and use of tools, and in the creation of social taboos and rituals. Caution must always be exercised in making direct comparisons between animals and people, and most of these areas are only beginning to be studied in any detail. Nevertheless, primate (homo)sexuality and the "traditions" associated with it offer us a striking mirror of some of our most "human" characteristics — and perhaps even a window into our evolutionary past and cultural history.
Language
Bonobos (also sometimes known as Pygmy Chimpanzees) have one of the most varied sexual repertoires of any species, with a wide variety of behaviors and positions used in sexual interactions, both heterosexual and homosexual. As a result, some Bonobos have developed an extraordinary system of gestural communication that is used specifically during sex. First discovered by pioneering ape-language researcher Susan Savage-Rumbaugh and her colleagues in the mid 1970s, this gestural system has far-reaching implications for our understanding of primate communication systems and the development of human language.53
A "lexicon" of about a dozen hand and arm gestures — each with a specific meaning — is utilized by Bonobos to initiate sexual activity and negotiate various body positions with a partner (of the same or opposite sex). For example, one gesture involves flicking the hand back-and-forth sideways from the wrist, meaning (approximately), "Move your genitals around"; this is used to get one's partner to position his or her genitals so as to facilitate a sexual interaction. Another gesture, lifting the arm with the palm downward, is employed when a Bonobo wants a sexual partner to move into a facing position for copulation. A catalog of some of the other gestures is given in the accompanying illustration. Hand signals may even be strung together in short sequences, and there is some evidence that the order of the gestures is significant.54 These manual signs are used during both heterosexual and homosexual activity, although it appears that they may be more prevalent in opposite-sex interactions, and males and females may also employ some of the gestures with different frequencies.
Most of the hand signals are
iconic, which means that they bear a physical resemblance to the meanings (i.e., body movements) they represent, although some are less transparently representational than others. Thus, the meaning "Turn your body around" can be related to the circular movement of the corresponding gesture, but this meaning is also conventionalized, since the intended action conveyed by the gesture must be agreed upon and understood by both participants for the gesture to be effective. An individual who hasn't learned the gestural system, for instance, wouldn't necessarily know what sort of "turning around" is intended by such a gesture. In conjunction with these hand gestures, a variety of positioning movements involving direct touching and placement of a partner's body or limbs are also used to facilitate sexual interactions. Together with the more abstract manual gestures, a total of at least 25 signals used during sex have been identified. In addition, patterns of eye contact and gaze also appear to have significant communicative value.
{67}

DESCRIPTION:
Arm is partially extended, hand flicks back and forth with sideways movement from wrist
MEANING:
"Move your genitals around" — used to get partner to position his or her genitals to facilitate a sexual interaction

DESCRIPTION: Arm is extended, hand bends at wrist and makes rapid, vigorous circular motions
MEANING: "Turn your body around" — also used as an invitation for a sexual interaction if other methods of initiation have not worked

DESCRIPTION: Hand is held outstretched toward partner (arm extends upward and outward with palm of hand angled toward the other individual)
MEANING: "Approach" — used to get partner to move closer; also used simply as an invitation for a sexual interaction
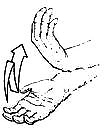
DESCRIPTION: Arm is extended, hand bends at wrist and flips toward self
MEANING: "Come here" — used to get partner to move closer during a sexual interaction

DESCRIPTION: Arm is partially extended and raised while hand flips upward at wrist
MEANING: "Stand up" — used to get partner to stand on hind legs before engaging in a face-to-face sexual interaction

DESCRIPTION: Arm is extended and raised till about head level with palm facing downward and placed lightly on another individual's back or shoulder
MEANING: An invitation for a sexual interaction

DESCRIPTION: Knuckles are rested on partner's arm or back and arm moves back toward self
MEANING: "Move closer" — used to get partner to assume a stance compatible with front-to-back sexual interaction

DESCRIPTION: Arm is lifted up with palm down
MEANING: "Position yourself" — used to get partner to move into a face-to-face position

DESCRIPTION: Hand and forearm move across body in a sweeping motion
MEANING: "Turn around" — used to get partner to rotate his or her entire body
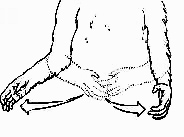
DESCRIPTION: Both arms wave or open out from body
MEANING:
"Spread your legs and/or arms" — used to get partner to open limbs to facilitate
face-to-face positioning
{68}
Researchers studying the Bonobo gestural system have suggested that the more abstract hand signals may have developed out of the simpler positioning movements.55 Communication during sex may initially have involved only fairly crude attempts to move one's partner, from which more ritualized touching and directive gestures may have evolved, which then gradually became more and more stylized until, in some cases, fairly opaque hand gestures resulted. This sequence is significant, because it represents the sort of progression from purely representational gestures to highly codified manual signals that has been identified in the development of human sign languages.56 More broadly, it shows the beginnings of abstraction or arbitrariness — that is, the creation of
symbols — which is a hallmark of human language in general. The Bonobo gestural repertoire is certainly not a "language" in the true sense of a complete human linguistic system, and by no means does it have the complexity or subtlety of even the simplest human gestural systems, let alone of a fully developed signed, spoken, or written language. Nevertheless, it is a formalized communication system that exhibits a level of sophistication unparalleled in any other nonhuman primate — perhaps even with a rudimentary "syntax" (in terms of the ordering of gestures) — and it may indeed represent a precursor to human language.
Even more significantly, Bonobos devised this system spontaneously: they invented the hand signals on their own and were not taught to use them by people. Attempts to teach apes various forms of human sign language or other communication systems have demonstrated that our primate relatives in many instances possess formidable linguistic capacities, but in all cases human prompting and intervention (at least initially) are involved. What is unique about the sexual gestures is that Bonobos themselves developed the hand signals and taught them to each other (or learned them from one another), in response to a communicative need that arose naturally within their own social interactions.57 Moreover, the specific social context that prompted this development is also unique: it was sexual behavior, or rather, the highly variable and plastic nature of Bonobo sexuality, that led to this development. Because of the wide variety of heterosexual and homosexual activities that characterize Bonobo sexual interactions, a supplementary communication system arose to help negotiate sexual interactions. In response to an unsurpassed sexual capacity in this species — including prominent homosexual activities — an unsurpassed animal communication system was created.
Not only is the Bonobo gestural system an outstanding example of the spontaneous development of cultural traditions in animals, it offers some clues into the origin of the human linguistic capacity. A number of theorists have suggested that the first human communication system may indeed have been a gestural language, that is, a system of hand signals.58 Why language should have evolved in the first place among human beings, however, is a subject shrouded in mystery and controversy. Of the many theories that have been put forward, a number suggest that language developed in response to social factors, such as the need to coordinate complex group activities such as hunting or farming. The Bonobo system demonstrates that another factor may also have been involved — one that is rarely, if ever, considered in discussions of the origin of language: sexuality. In particular, as {69}
sexual interactions became more variable over the course of evolution, gestural systems of greater complexity may have developed to facilitate sexual encounters.
Primate evolution has been characterized by an ever-increasing separation of sexuality from its reproductive "functions," including the development of numerous types of homosexual and nonprocreative heterosexual activities — most prominent in human beings and Bonobos (considered by some to be the primate most similar to humans), less so in Common Chimpanzees, Gorillas, and other apes. Scientists have also identified a corresponding increase in complexity of communication systems used during sexual interactions among apes, proceeding from Gorillas to Common Chimpanzees to Bonobos (and onward, of course, to human beings).59 The progression is probably not quite as orderly as this sequence suggests, and multiple factors are surely involved in the genesis of each species' particular communication systems. The general trend, however, is clear: as sexual interactions become more variable, sexual communication systems become more sophisticated. It is possible, therefore, that sexuality — particularly the fluidity associated with nonreproductive sexual practices — played a significant role in the origin and development of human language.
Tools
A hallmark of human cultural evolution was the development of tools, later elaborated into the full array of material technologies that we know today. But many animals, especially primates, also use inanimate objects to manipulate or affect things in their environment in ways that can be seen as precursors to similar activities in human beings. Over 20 different types of tool use have been identified in primates and other species — Common Chimpanzees, for example, employ objects as weapons, as levers, and as drinking and feeding implements of various sorts (such as the well-known example of sticks being used to capture and eat termites or ants). Tools can also be used to affect an animal's own (or another's) body, for example as part of a "hygiene" or grooming regimen. Chimpanzees and other primates, for example, often use leaves, twigs, straw, rags, or other objects to clean themselves and wipe away bodily secretions (such as saliva, blood, semen, feces, and urine). Chimps and Savanna Baboons have also been observed using sticks, twigs, and stones to clean their own or each other's teeth (and even perform dental extractions). Chimps also sometimes tickle themselves with various items such as stones or sticks, and Japanese Macaques occasionally use similar items to groom one another.60
Less well known, however, is the use of objects for purposes of sexual stimulation. A number of primates employ various implements as masturbatory aids (in both the wild and captivity), and this aspect of tool culture has not received widespread attention in discussions of the development of animal and human object manipulation. Female Orang-utans, for instance, sometimes masturbate by rubbing objects on the clitoris or inserting them into the vagina; tools used for this purpose include pieces of liana bitten off to an appropriate size or (in captivity) pieces of wire. Male Orangs also use objects to stimulate their genitals, including {70}
one individual who ingeniously fashioned an implement by pushing a hole through a leaf with his finger. He inserted his erect penis into this "orifice," then rubbed the leaf up and down the shaft to stimulate himself. Males also sometimes hold a piece of fruit (such as an orange peel) in their hand and masturbate against it.61
Common Chimpanzees have also developed several innovative masturbation techniques using a variety of different tools. One female gathered a small collection of sticks, pebbles, and leaves, from which she would carefully select a particular item to stimulate herself with. By placing a leaf underneath her vulva, for example, and flicking the stem with her knuckle, she made the leaf vibrate and thereby externally stimulate her genitals. She also repeatedly inserted the stem into her vagina, often lubricating it with saliva and manipulating it with her hand so as to stimulate herself internally. In one instance, she rocked back and forth with the stem inserted, rubbing the leaf against a vertical surface so that the stem actually vibrated inside her. On other occasions, she repeatedly inserted and withdrew a pebble from her vagina or used a small stick to stimulate her genitals. Other female Chimps have also been observed rubbing or tickling their external genitals with items or inserting them into the vagina, including pieces of mango, twigs, and leaves, as well as man-made objects such as small boxes or balls. Similarly, several young males assembled collections of stones, fruits, or even pieces of dried dung, which they would thrust against to stimulate their genitals. Male and female Bonobos occasionally employ inanimate objects for masturbation as well, stimulating themselves with (or thrusting against) branches, wood shavings, and other items.62
Like Orang-utans, one female Bonnet Macaque invented some relatively sophisticated techniques of tool manufacture, regularly employing five specific methods to create or modify natural objects for insertion into her vagina. For example, she stripped dry eucalyptus leaves of their foliage with her fingers or teeth and then broke the midrib into a piece less than an inch long. She also slit dry acacia leaves in half lengthwise (using only a single half) and fashioned short sticks by breaking {71}
longer ones into several pieces or detaching portions of a branch. Implements were also sometimes vigorously rubbed with her fingers or between her palms prior to being inserted into her vagina, and twigs, leaves, or grass blades were occasionally used unmodified.63
The use and manufacture of tools by primates is considered an important example of cultural behavior in animals, and a forerunner of the activities that are so widespread among human beings. Although many different forms and functions are evident in animal tool use, these examples show that nonreproductive sexual activities are part of the overall behavioral pattern: the primate capacity for object manipulation extends seamlessly into the sexual sphere. Apes and monkeys use a variety of objects to masturbate with and even deliberately create implements for sexual stimulation by cutting or forming materials such as leaves or twigs (often in highly creative ways). Similar types of activities occur among people, of course, and sexual implements of various sorts have a long and distinguished history in human culture. Dildos or phalli made of stone, terra-cotta, wood, or leather, for example, were used in ceremonial "deflowering" and fertility rites — as well as for masturbation and inducing sexual pleasure in a partner — in ancient Egypt, Rome, Greece, India, Japan, and Europe. Examples have been recorded from at least as far back as the Paleolithic through medieval times — including some biblical references — as well as in the ongoing traditions of many indigenous peoples throughout the world.64 However, few (if any) anthropologists have ever considered the possibility that sexual stimulation may have been a component of tool use among early humans or even played a part in the origin and elaboration of material culture. Of course, technological complexity is not the only measure of cultural development — some of the most complex linguistic and oral history traditions, for example, are to be found among the South African San peoples and the Australian Aborigines, whose material culture is relatively simple. And certainly many more "utilitarian" functions can be identified in the development of tool use among our human, protohuman, and primate ancestors. Nevertheless, the pursuit of sexual pleasure may have contributed, in some measure, to our own heritage as creatures whose tool-using practices are among the most polymorphous of any primate.
Taboo
The vast majority of human cultures prohibit sexual relations between people who are related. There is still ongoing debate among scientists as to whether this prohibition — commonly known as the incest taboo — is instinctual or learned. Regardless of the extent to which biological factors are involved, there are clearly strong social and cultural components to incest avoidance. Different human cultures and societies vary widely in how they define incestuous relations and to what extent such activities are both stigmatized and practiced. For example, although parental incest (father-daughter, mother-son) is prohibited in virtually all societies (yet still occurs, despite such prohibition, with varying frequencies), there is wider latitude regarding other blood relations. Cousin marriage is considered acceptable in some cases, unacceptable in others, while some societies make a further distinction {72}
between relations with cross cousins as opposed to parallel cousins — a biologically arbitrary distinction, since there is no evidence of any greater genetic "harm" in one form of cousin marriage than another. Brother-sister marriage was widely practiced in ancient Roman Egypt, and among the royal families of some central African and Balinese societies, ancient Incans, Hawaiians, Iranians, and Egyptians — in fact, Cleopatra is thought to have been the product of 11 generations of incestuous marriages within the Ptolemaic dynasty.65
Further evidence of a learned or cultural component to incest prohibitions relates to the role played by social familiarity as opposed to genetic relatedness in choice of partners. In our culture, sexual relations between adoptive or stepfamily members are generally frowned upon even though the individuals involved are not related by blood. Conversely, people who are genetically related but, because of social circumstance (e.g., separation at birth), are unaware of their biological connection may develop a relationship (at least until they learn of their relatedness). Other societies vary considerably in this regard: in the Israeli kibbutzim, for instance, unrelated individuals who are brought up together hardly ever marry one another. In contrast, a traditional form of Taiwanese marriage involved girls being adopted into families as children and then, on reaching adulthood, marrying their stepbrothers, although such marriages were considered less preferable than other arrangements. Among the Arapesh people of New Guinea, a similar practice of stepsister marriage was widely accepted and preferred.
The fact that homosexual relations are usually prohibited between related individuals also points to the importance of nonbiological factors in the incest taboo. In most human cultures that "permit" some form of same-sex eroticism, from contemporary America to indigenous tribes of New Guinea, the choice of homosexual partners is subject to distinctions of "kin" and "nonkin." This is in spite of the fact that no children, and hence no potentially harmful genetic effects, can result from such unions. Typically the same restrictions are applied to homosexual as to heterosexual relations. In a number of New Guinean societies, however, slightly different kinship constraints regulate the choice of same-sex and opposite-sex partners. In fact, homosexual partners in some tribes are actually required to be
more distantly related than heterosexual ones — the exact opposite of what would be expected if incest taboos were based solely on biological factors.66
This is significant, because most theories about the biological basis of the incest taboo focus on the potential for increased rates of birth defects and lower genetic variability as a result of inbreeding. Even for heterosexual relations, though, the evidence is not nearly as unequivocal as one would suppose: numerous studies of small populations that have practiced inbreeding for many generations reveal no deleterious effects, owing to the rapid elimination of genetic defects and subsequent stabilization of the gene pool.67 To adduce further evidence for a biological basis to the prohibition, scientists often point to the existence of "incest taboos" in animals. Ironically, though, many animal species actually show evidence of a "cultural" or "social" dimension to their avoidance of sexual activity between relatives that parallels the human examples — most notably among primates, and most notably involving homosexuality.
{73}
There is a great diversity of incestuous activity among animals, not only in the frequency and types of relations that occur but also in the degree to which such activity is avoided or pursued, in both heterosexual and homosexual contexts. Even among primates, many different scenarios and versions of "taboos" are found. In Rhesus Macaques, for instance, incest of any sort is not common although mother-son, brother-sister, and brother-brother relations do occur (some males actually appear to prefer mating with their mothers). In Gibbons, heterosexual incest (both parental and sibling) is sometimes practiced and homosexual relations are almost always incestuous, while both heterosexual and homosexual activity between siblings (or half-siblings) occurs in Gorillas. Most strikingly, several species appear to have developed systematic homosexual "incest taboos," each with its own socially defined set of "acceptable" and "unacceptable" partners. In some cases, these restrictions differ significantly from those governing the corresponding heterosexual relations (as in some human populations).
Homosexual consortships (pair-bonding with sexual activity) among Japanese Macaques, for example, virtually never occur between mothers and daughters or sisters. In contrast, heterosexual brother-sister or mother-son relations, while not common, are much more prevalent than homosexual incest in this species. Interestingly, aunts and nieces among Japanese Macaques do not generally recognize each other as kin — when intervening on behalf of individuals during aggressive encounters, for example, aunts do not assist their nieces any more often than do nonrelatives, and significantly less often than do mothers, grandmothers, and sisters. Consequently, some blood relatives are able to form consortships with one another: about a third of all aunt-niece dyads interact homosexually. In other words, Japanese Macaques have an overall pattern of incest avoidance unique to homosexual relations, within which apparently incestuous aunt-niece pairs are "permitted" because such partners do not count as kin in the larger social framework.
In Hanuman Langurs, both heterosexual and homosexual incest taboos are in effect, but with slightly different restrictions. Heterosexual incestuous relations of any kind are generally avoided; sexual activity between mothers and daughters is also "prohibited" (accounting for only about 1 percent of all homosexual mounting). In contrast, half sisters (females with the same mother but different fathers) are "allowed" to have sexual relations with one another — and in fact, more than a quarter of all mounts between females occur between half sisters. In Bonobos, incestuous relations between females also generally appear to be avoided: when females immigrate into new troops as adolescents, they are usually unrelated to most of the other troop members, but sexual activity is not practiced with those females to whom they are related.68
These examples demonstrate that, at least in some nonhuman primates, homosexual (as well as heterosexual) relations are subject to various social prohibitions regarding choice of appropriate partners, especially where relatives are involved. These choices are not due to instinct (i.e., avoidance of an activity that would yield harmful genetic effects), because no offspring result from such relations, and because not all incest taboos are identical. Nor are they simply a "carryover" from heterosexual taboos, because same-sex and opposite-sex relations often have different {74}
prohibitions. Crucial distinctions exist between species, populations, and even between heterosexual and homosexual activity in the same species, concerning "allowable" incestuous relations — differences that cannot be attributed solely to biological (genetic) factors. Only some relatives actually "count" as related for the purpose of incest taboos, and which individuals are "tabooed" is, to a large extent, arbitrary. In other words, primates must
learn what sort of kinship system(s), if any, govern sexual associations — both homosexual and heterosexual — in their social group. While the occurrence and expression of same-sex activity in these animals very likely has an instinctual or genetic component as well, homosexual relations exhibit important "cultural" characteristics that probably involve a high degree of social learning. "Taboos" exist in animals, and homosexuality is one area where such prohibitions manifest themselves in particularly compelling ways.
Ritual
Where do human rituals such as taking an oath come from? In a fascinating study of Savanna Baboon social systems, primatologists Barbara Smuts and John Watanabe offer a startling answer: they suggest that such symbolic gestures might be traced to the ritualized homosexual activities that take place between male Baboons.
69 As part of their social interactions, male Savanna Baboons perform a variety of formalized sexual and affectionate behaviors with each other, most notably "diddling," that is, fondling of the penis and scrotum. Other ritualized homosexual activities include mounting; grabbing, fingering, and nuzzling of the rump; kissing and nuzzling of the genitals; and embracing and kissing on the head or mouth {75}
(similar activities are also found in a number of other primates, including Common Chimpanzees, Bonnet Macaques, and Crested Black Macaques). Although these behaviors undoubtedly have an affectionate or "pleasurable" tactile component as well as a sexual dimension, they have also been characterized by some scientists as "greetings" interactions, and it is thought that they may serve to negotiate and solidify cooperation between males as well. Indeed, two males sometimes pair up and form a stable "coalition" with each other in which their mutual defense and aid is symbolized by the reciprocity of their ritual sexual exchanges with one another. Smuts and Watanabe suggest that sexual gestures such as diddling, which involve one male placing his most vulnerable and intimate organs literally in the hands of another, are in a sense a prototypical form of oath-swearing: one male, by his actions, is indicating his trust and commitment to cooperate with the other.
But what does this have to do with human rituals of oath-taking? In our society at least, oaths usually involve gestures such as raising of the right hand, crossing the heart, or perhaps even placing the hand on a Bible, but certainly nothing so forward as fondling of the genitals. Surprising as it may seem, though, Smuts and Watanabe present some intriguing clues that gestures similar to the ritual homosexual activities of Baboons (and other primates) may in fact have been a part of human oath-taking at one time and are even still used in some contemporary cultures. In a number of Australian Aboriginal tribes, for example, holding of the penis is traditionally used as a gesture to express male allegiance and cooperation, as well as a ritual part of resolving disputes between "accused" and "defending" parties. Among the Walbiri and Aranda people, when different communities get together or when grievances need to be settled in formal "trials," men participate in what is known variously as touch-penis, penis-offering, or the penis-holding rite. Each man presents his semi-erect organ to all the others in turn, pressing it into each man's palm and drawing it along the length of the upturned hand (held with the fingers toward the testicles). By offering and grasping each other's penis — said to represent "paying with one's life" — the men make an avowal of mutual support and goodwill between them, or symbolize and solidify the agreement they have reached during the settling of a dispute. A similar gesture involving stroking of the genitals and/or scrotum is used as a greeting in some New Guinea tribes such as the Eipo and Bedamini.70
Closer to home, there is historical — even biblical — evidence that similar rituals may have been a part of the Judeo-Christian and Euro-American heritage. Ironically, the book that is today used in so many of our own oath-swearing ceremonies contains within it an allusion to these earlier practices. In Genesis 24:9 there is a reference to the servant of Abraham swearing an oath by placing his hand under his master's "loins." Moreover, according to the
Oxford English Dictionary the words testify, testimony, and
testicle are probably all related, sharing the common root testis, which originally meant "witness." Although these connections are somewhat speculative, they suggest a line of continuity between ritualized homosexual behavior in primates and human social rituals such as oath-taking. As Smuts and Watanabe point out, notions of truth and sanctity as expressed by the human ceremonies are vastly different from those of Baboons (if present at all in the nonhuman {76}
context). Nevertheless, the
forms of these rituals — and their social outcomes — are strikingly similar.
Language, tools, taboo, ritual — each of these is part of a larger puzzle or
matrix of cultural development that is traditionally seen as distinct and distant from sexuality. Nevertheless, the occurrence of a number of remarkable primate behaviors revolving around homosexuality and nonreproductive heterosexuality suggests that these domains are much more intimately associated than previously imagined. Sexual gesture systems, masturbatory tools, homosexual incest taboos, and ritualized same-sex "oath-taking" offer extraordinary juxtapositions of culture, biology, society, and evolution. Primate (homo)sexual behaviors such as these exemplify both cultural traditions and evolutionary inheritances. In turn, they may have contributed to the development of some of the most hallowed and cherished landmarks of human cultural history as well.71
Unnatural Nature
Animals don't do it, so why should we? Can you even imagine a queer grizzly bear? Or a lesbian owl or salmon?
— from a letter written to Dean Hamer, coauthor of The Science of Desire: The Search for the Gay Gene and the Biology of Behavior72
Many people, such as the man quoted above, believe that homosexuality does not occur in nature and use this belief to justify their opinions about human homosexuality. In fact, rarely is homosexuality in animals discussed on its own: inevitably, cross-species comparisons are drawn to ascribe moral value to the behavior — both positive and negative. Nowhere is this more apparent than in the notion of "naturalness" and the entire complex of animal-human comparisons that this problematic term evokes. The prevailing view is an overly simplistic one: if homosexuality is believed to occur in animals, it is considered to be "natural" and therefore acceptable in humans; if it is thought not to occur in animals, it is considered "unnatural" and therefore unacceptable in humans. The debate seems clear and the lines of distinction inviolable.
Any careful consideration of the logic behind the equation
occurs in animals = natural = acceptable in humans will show, however, that this line of reasoning is flawed. As many people have pointed out, humans engage in a wide variety of behaviors that do not occur in nature, from cooking to writing letters to wearing clothes, and yet we do not condemn these activities as "unnatural" because they are not found among animals. As author Jon Ward explains, with regard to a friend who asserted "You can't argue with biology" (believing that homosexuality was "unnatural"):
«Has he never fried an egg? The whole of human history is an "argument with biology." The very civilization which the most homophobic ideologues are eager to defend is the antithesis of nature: law and art.»73
{77}
We also use our biology and anatomy in ways that "nature did not intend for them to be used" without ascribing a moral value to such activities. As James Weinrich observes, the tongue's primary biological purpose is for the act of eating, yet its use in acts of speech, bubble-gum-blowing, or kissing is not therefore considered "unnatural." In addition, many things that do occur spontaneously in animals — diseases, birth defects, rape, and cannibalism, for example — are not considered to be "natural" or desirable conditions or behaviors in most humans. Weinrich aptly remarks, "When animals do something that we like, we call it natural. When they do something that we don't like, we call it animalistic."74
The Natural History of Homosexuality
The historical record also shows that attitudes toward homosexuality have little to do with whether people believe it occurs in animals or not, and consequently, in its "naturalness." True, throughout much of recorded history, the charge of "unnaturalness" — including the claim that homosexuality did not occur in animals — was used to justify every imaginable form of sanction, control, and repression against homosexuality. But many other interpretations of "naturalness" were also prevalent at various times. Indeed, the very fact that homosexuality was thought to be "unnatural" — that is, not found in nature — was sometimes used to justify its
superiority to heterosexuality. In ancient Greece, for example, same-sex love was thought to be purer than opposite-sex love because it did not involve procreation or "animal-like" passions. On the other hand, homosexuality was sometimes condemned precisely because it was considered
closer to "nature," reflecting the base, uncontrolled sexual instincts of the animal world. The Nazis used this reasoning (in part) to target homosexuals and other "subhumans" for the concentration camps (where homosexual men subjected to medical experiments were referred to as "test animals"), while sexual relations between women were disparagingly characterized as "animal love" in late eighteenth-century New England. The irrationality of such beliefs is highlighted in cases where charges of "unnaturalness" were combined, paradoxically, with accusations of animalistic behavior. Some early Latin texts, for instance, simultaneously condemned homosexuals for exhibiting behavior unknown in animals while also denouncing them for imitating particular species (such as the hyena or hare) that were believed to indulge in homosexuality.75
In our own time, the fact that a given characteristic of a minority human population is biologically determined has little to do with whether that population should be — or is-discriminated against. Racial minorities, for example, can claim a biological basis for their difference, yet this has done little to eliminate racial prejudice. Religious groups, on the other hand, can claim no such biological prerogative, and yet this does not invalidate the entitlement of such groups to freedom from discrimination. It should be clear, then, that whether homosexuality is biologically determined or not, whether one chooses to be gay or is born that way, or whether homosexuality occurs in nature or not — none of these things guarantees the acceptance or rejection of homosexuality or in itself renders homosexuality "valid" or "illegitimate."
{78}
The debate about the "nature" and origin of homosexuality often invokes seemingly opposite categories: genetics versus environment, biology versus culture, nature versus nurture, essentialism versus constructionism. Indeed, the very categories "homosexual" and "heterosexual" are themselves examples of such a dichotomy. By using these categories, biologists and social scientists hope to discover what aspects of homosexuality, if any, are biologically determined. Yet by framing the debate in terms of such categories, it is easy to forget that more complex interactions between factors must be considered. For example, most research shows that both environment
and biology are relevant in determining sexual orientation in people (and probably also animals). Some individuals may have an innate predisposition for homosexuality, but the right combination of environmental (including social) factors is required for this to be realized. And how meaningful is it to talk about a culture-nature distinction when, as we have just seen, some animal species have themselves developed forms of cultural behavior? Similarly, by focusing attention on the "causes" of homosexuality, the determinants of heterosexuality are considered irrelevant — or, alternatively, heterosexuality is assumed to be inevitable unless something "goes wrong." And not all sexuality fits neatly into the categories of exclusive homosexuality or exclusive heterosexuality — the large realm of experience that involves bisexuality is easily glossed over in discussions about the origins of homosexuality/heterosexuality. So, too, with the question of whether homosexuality is "natural" and what its occurrence in animals can tell us about this: things are considerably more complicated than they initially appear.
What is remarkable about the entire debate about the naturalness of homosexuality is the frequent absence of any reference to concrete facts or accurate, comprehensive information about animal homosexuality. Those who argue against the naturalness of homosexuality assert with impunity that same-sex behavior does not occur in nature (like the man quoted above) and is therefore self-evidently abnormal. Those who argue in favor of a biological origin for homosexuality often ignore the complexities of animal behavior arising from social, protocultural, or individual life-history factors (relying on the behavior of laboratory animals injected with hormones, for example, instead of long-term studies of animals interacting in their own social groups or communities).76 This is because naturalness is more a matter of interpretation than facts. Now that the widespread occurrence of animal homosexuality is beginning to be documented, little if anything is likely to change in this discussion. More information about same-sex activity in animals simply means more possible interpretations: the information can be used to support or refute a variety of positions on the naturalness or acceptability of homosexuality, depending (as before) on the particular outlook of whoever is drawing the conclusions.
As James Weinrich points out, the only claim about naturalness that is actually consistent with the facts is the following: homosexual behavior is as natural as heterosexual behavior.77 What this means is that homosexuality is found in virtually all animal groups, in virtually all geographic areas and time periods, and in a wide variety of forms — as are heterosexuality, divorce, monogamy, and infanticide, among other things. Conversely, heterosexuality is as "unnatural" as homosexuality is, {79}
since it often exhibits social elaboration or cultural "embellishment," as well as many of the "unacceptable" features stereotypically associated with same-sex relations, such as promiscuity, nonreproduction, pursuit of sexual pleasure, and interactions marked by instability, ineptitude, and even hostility.78 But whether this means that homosexuality is "biologically determined" and/or "socially conditioned" — and by extension, (un)acceptable in humans — is largely a question of interpretation. Of course, from a scientific perspective, the sheer extent and variety of homosexual expression in the animal world reveals an aspect of nonhuman biology and social organization that is unexpected — one with far-reaching (perhaps even revolutionary) implications. It demands careful consideration and suggests a rethinking of some of our most fundamental notions of environment, culture, genetics, and evolutionary and social development. But to automatically conclude that because homosexuality occurs in animals, it must be biologically determined oversimplifies the debate and does an injustice to the facts.
For most people, animals are symbolic: their significance lies not in what they are, but in what we think they are. We ascribe meanings and values to their existence and behaviors in ways that usually have little to do with their biological and social realities, treating them as emblems of nature's purity or bestiality in order to justify, ultimately, our views of other human beings. The animals themselves remain enigmatic, mute in the face of this seemingly endless onslaught of human interpretations of their lives. If this were merely a matter of debate among people, it could perhaps be put in its proper perspective as simply yet another human folly. Unfortunately, the interpretations applied to animal (sexual) behaviors by people are far from innocuous: they can have grave consequences, or even be a matter of life or death — for both humans and animals alike. When a gay man or lesbian is assaulted or murdered because the attacker thinks that homosexuality is "unnatural," for example, or when politicians' legislative and judicial decisions concerning homosexuality are coded in such terms as "crimes against nature," much more is at stake than the scientific interpretation of animal behavior.79
The moral value ascribed to animal sexuality can also impact directly on the welfare of the creatures themselves. In 1995 a biologist with the U.S. Fish and Wildlife Service briefed Senator Jesse Helms's staff about the value of saving an endangered bird, the red-cockaded woodpecker, which lives in the southeastern United States.80 His presentation stressed the supposed "family values" of the species, referring to the birds' monogamous and relatively long-lasting heterosexual pair-bonds. In other words, the right of this species to exist — as determined by legislators voting on the Endangered Species Act — was predicated not on its intrinsic value, but on how closely its behavior could be made to resemble what is currently considered acceptable conduct for humans. And this is most definitely a case of presenting an idealized "image" of the species: the red-cockaded woodpecker's "family values" are in reality far more complex, messy, and "questionable" than what the politicians were told.
True, this species usually breeds in long-term, monogamous pairs, but its social life is replete with variations on this theme, some of which Senator Helms would have found downright horrifying.81 Many family groups in this species are unstable: {80}
one six-year investigation found that only six out of thirteen breeding pairs remained together, while studies of the species in Helms's home state of North Carolina revealed that nearly 20 percent of females in this population desert their mates and switch family groups. Males sometimes leave their partners as well, and the overall (species-wide) divorce rate is about 5 percent; nonmonogamous copulations also occasionally occur, with slightly more than 1 percent of nestlings being fathered by a male other than their mother's mate. Red-cockaded woodpeckers also frequently live in "stepfamilies" or "blended families": more than a quarter of the younger birds who live in breeding groups and help with parenting duties may be related to only one parent, and 5-11 percent are related to neither. Some of these "helper" birds engage in decidedly un-family-like activities, such as ousting a parent from its group or even committing "stepfamily incest" by mating with the remaining parent. Incest involving full or half siblings, though rare, also occurs. Other helpers forgo reproduction entirely (continuing to live with their parents as adults for several years), and there are also solitary nonbreeding birds in the population, as well as all-male groups. Some red-cockaded woodpecker groups may also be polygamous or "plural" breeding units, with two females both breeding (or trying to breed) at the same time.
Would the red-cockaded woodpecker be considered less "deserving" of protection if Senator Helms and his staff learned that these birds participate in nonreproductive sexual activity (mating during incubation, or long before egg-laying), or siblicide and starving of offspring, or infanticide and chick-tossing from the nest? All of these behaviors have been documented in this species, yet none were included in the scientific presentation to the politicians in whose hands this bird's fate rests, for they would shatter the illusion of its "family values." Homosexual activity has not (yet) been observed in red-cockaded woodpeckers, although it does occur in related species such as Acorn Woodpeckers and Black-rumped Flamebacks. Should such behavior come to light, one can only dread the consequences for this, or any other, endangered species whose survival depends on human assessment of its "moral conduct."
Homosexuality has a "natural history" in every sense of the term: that is, it has both biological ("natural") and social or cultural ("historical") dimensions that are interconnected and inseparable. It is not a uniform phenomenon in either animals or people: it takes many forms, and it exhibits numerous variations and idiosyncrasies. The interplay of biology and environment in shaping these features — and indeed, the very definitions of what is "cultural" as opposed to "biological" — is far more complex than polarized debates would have us believe. Because the discussion is often framed in terms of misleading dichotomies such as "nature versus nurture" or "genetics versus environment," the possibility that both are relevant (and can influence one another) is repeatedly overlooked, as is the possibility that sexual behavior in some animals has a significant sociocultural component. Yes, homosexuality occurs in nature and apparently always has. But does this make it "natural" or simply "animalistic"? The answer to this question is entirely in the eye of the beholder, rather than in any inherent quality or context of the phenomenon itself.
{81}
Homosexuality in the Home and on the Farm: Pets and Domesticated Animals
Nowhere are questions of interpretation more vividly illustrated than with the animals we often consider to be the most "human" — our pets. Here it is not genetic similarity (as with primates) but our emotional and physical proximity that establishes the animal-human connection, whether it is with the creatures we keep as companions or domesticated animals on farms and ranches. Same-sex activity is readily familiar to pet owners and animal trainers/handlers, many of whom can give examples of homosexual mounting, pair-bonding, or other same-sex (or bisexual) activities in their own animals or those of friends. These anecdotal reports have been confirmed by scientific studies of domesticated animals.82 Same-sex pair-bonding as well as homosexual mounting (including ejaculation during interactions between males) has been documented in Dogs, including breeds such Beagles, Basenjis, Cocker Spaniels, and Weimaraners. Examples of both female and male homosexual behavior in Cats have also been confirmed, including mutual genital stimulation and mounting between females, and mounting leading to orgasm among males. Homosexuality has also been verified in other animals kept as pets, including same-sex courtship and mounting in Guinea Pigs; homosexual mounting in female domesticated Rabbits and Hamsters; and same-sex pair-bonding, courtship, and mounting among caged birds such as Zebra Finches, Bengalese Finches, and Budgerigars. Many common aquarium fish also exhibit homosexuality or transsexuality.
Homosexual behavior has been studied in a wide variety of domesticated and farm animals as well: Cattle, Sheep, Goats, Pigs, and Horses of both sexes participate in homosexual mounting, while same-sex pair-bonds have also been reported for Pigs, Sheep, and Goats. In fact, homosexual activity is so routine among domesticated hoofed mammals that farmers and animal breeders have coined special terms for such behavior: mounting among male Cattle is referred to as the buller syndrome (steers who are mounted are called bullers, the males who mount them are riders), female sows who mount each other are described as "going boaring," mares who do so are said to "horse," while cows are said to "bull." Same-sex activity is often utilized, paradoxically, in breeding programs. In some species homosexual mounting among females is used as a reliable indicator of when they are in heat, while young bulls or steers (known as teasers) are often presented to mature bulls to arouse them and allow their semen to be collected (for later use in artificial insemination) .83
For each domesticated species that exhibits homosexual behavior, there are one or more wild "relatives" in which homosexuality has also been observed: Lions and other wild Cats, Wolves and other wild Dogs, Cavies (the wild ancestors of Guinea Pigs), American Bison and other wild Buffalo, Bighorn Sheep and other wild Sheep and Goat species, Zebra and other wild Horses, and so on. In some cases, the same-sex activities observed in domesticated species and their wild ancestors exhibit striking similarities: the group sexual interactions or "huddles" of Goats and Sheep (both wild and domesticated), the frequency of same-sex mounting in domestic {82}
Cattle and wild Buffalo, female Cats and Lions placing themselves underneath a female partner to invite mounting, or the same-sex courtship displays by female Turkeys and wild Sage Grouse. In other cases, there are equally striking dissimilarities: pair-bonding and mounting among domesticated boars, for instance, contrasted with the virtual absence of same-sex activity in male wild Pigs; or fairly extensive homosexual courtship activities in female Cheetahs, contrasted with little, if any, such courtship activities in domestic Cats.
Although scientifically verified, homosexuality in pets and other domesticated animals continues to evoke many meanings for the people who simply live or work with such creatures, independent of the "facts." As with all human observations of the animal world, people tend to see only what they are prepared to accept. This is illustrated quite clearly in two contrasting views of homosexual behavior in farm animals, symbolizing the contradictory interpretations of same-sex activity that are applied to both people and animals. Anita Bryant, in a particularly brilliant turn of logic, once asserted that "even barnyard animals don't do what homosexuals do." When informed that barnyard animals and many wild species actually do "stoop" to the level of human homosexuals, she retorted, "That still doesn't make it right."84 Not surprisingly, noted lesbian author and historian Lillian Faderman offers a markedly different view:
«It's ridiculous for people not to recognize it [homosexuality] in nature. My partner once had a ranch, and I was just fascinated with the way the female animals would often mount other female animals as well as be mounted ... . Mammals are simply sexual.»85
Each of these women has strong opinions about animal homosexuality, and each woman's viewpoint is informed by her feelings about homosexuality in people. Of the two, however, Lillian Faderman's perspective is closer to the scientific reality of homosexuality in the animal kingdom. The next chapter explores in more detail the way that people have interpreted animal homosexuality throughout the history of science. Unfortunately, biologists themselves have often espoused views that have more in common with Anita Bryant's than Lillian Faderman's.
<<< ||||| >>>