<<< >>>
{566}
Perching Birds and Songbirds
COTINGAS, MANAKINS, AND OTHERS
 Cotingas
Cotingas
GUIANAN COCK-OF-THE-ROCK (Rupicola rupicola)
IDENTIFICATION: A small (10 inch) perching bird; adult males are brilliant orange with elaborate fringed wing plumes and an imposing helmetlike feather crest; adolescent males have mottled brown and orange plumage, while females are uniformly dark. DISTRIBUTION: Northern South America, primarily in southern Venezuela, the Guianas, adjacent parts of Brazil. HABITAT: Forests, usually near cliffs, mountains, or rock outcrops (on which females build their nests, giving the bird its name). STUDY AREA: Raleigh falls-Voltzberg Nature Reserve, Suriname, South America.
Social Organization
The spectacularly plumed Guianan Cock-of-the-Rock has what is known as a LEK social and mating system: males inhabit individual territories, usually clustered in the same area, which are used for display and courtship. Each display "court" consists of a cleared area on the forest floor and surrounding perches. Territories are maintained year-round, but courtship and mating occur only from late December through April. Females (and in this species, young males) visit these territories to choose which males they want to mate with. Other than this, males and females lead virtually separate lives: males do not participate in any aspect of nesting or parental care and rarely encounter females outside of the breeding season.

A younger (adolescent) male Guianan Cock-of-the-Rock mounting a bright orange adult male in the forests of Suriname
{567}
Description
Behavioral Expression: Homosexual activity between adult and adolescent males, as well as among adolescent males, is a routine occurrence in Guianan Cock-of-the-Rock. Males court and display to each other, as well as engage in homosexual mounting. A typical homosexual encounter begins when the adult males are perched beside their display courts, each glowing like an orange beacon in the jungle gloom. Both females and adolescent (yearling) males are attracted to the lek, which is carefully situated in the forest to take advantage of the ambient light characteristics of the environment (thereby showing off its owner to his best advantage). The courtship sequence commences with a GREETING DISPLAY: the males begin calling raucously, then each drops to the ground with a thump and begins beating his wings violently, flashing patches of black and white. This often produces a whistling sound as air rushes through the specially modified wing feathers.
This attention-grabbing sequence is followed by the GROUND DISPLAY: each adult male crouches down, fanning out the delicate filaments of his wing coverts, puffing out his chest and rump feathers and erecting his crest, resulting in a spectacular visual effect. By this time an adolescent male who is attracted by the display has landed beside the adult and is hopping about the display court, often crouching in a version of the courtship posture himself. The adult keeps his back toward the younger male at all times but is otherwise motionless, showing off his plumage to its best and inviting the adolescent to mount him. During homosexual copulation, the younger male climbs onto the adult's back and perches firmly, moving his tail sideways to try to make genital contact. Often the younger male mounts the older male several times in succession, and courtship and display often alternate with mountings. Sometimes males also mount each other in the trees surrounding a display court. Homosexual interactions differ from heterosexual ones in that both participants perform some version of the ground display; also, unlike females, neither male pecks at or touches the other's rump prior to mounting.
Adolescent males usually visit the display courts of several adult males, although some adults are clearly more "popular" than others because they receive more attention from the younger males. Typically, a yearling has homosexual interactions with anywhere from one to seven different adult males during the mating season. Nor does homosexual activity always involve an adult territory owner and a yearling male: adolescents often mount nonbreeding males who do not have their own display territories and sometimes also mount other adolescent males. Homosexual activity is not separate from heterosexual courtship and copulation, {568}
but takes place in the same locations and often while male-female interactions are happening in the vicinity. However, homosexual courtship and mounting often take priority over heterosexual interactions. If an adolescent male approaches an adult who is courting a female, the female usually leaves (or is chased away), and the two males turn their attentions to each other. Moreover, if a female encounters a male who is courting or involved sexually with another male, she usually waits until the adolescent male leaves before approaching the adult.
Frequency: Homosexual activity is very common among Guianan Cock-of-the-Rock: in fact, mountings between males are as frequent as male-female mountings, accounting for nearly half of all copulations. About 10 percent of heterosexual courtships are interrupted by adolescent males visiting the courting male; roughly one out of five of these "interruptions" involves courtship or sexual behavior between males. During the breeding season, homosexual activity may occur daily, and an adolescent male will usually have 6-7 homosexual encounters over the season (although some males engage in homosexual mounting more frequently, 15 or more times over a season).
Orientation: Close to 40 percent of the male population participates in some form of homosexual activity. Depending on his age, a bird that has same-sex interactions may or may not also engage in heterosexual activity. Among adult males (three years or older), nearly a quarter (23 percent) are mounted by other males, and 6 percent of these do not mate or court females at all. In fact, those adults who do not mate heterosexually are often the ones most frequently mounted by younger males. Among yearling males (virtually none of whom mount females), almost two-thirds (64 percent) engage in homosexual activity. Two-year-old males, on the other hand, very rarely engage in homosexual activity. Thus, when birds of all ages are taken into account, nearly 20 percent of the male population at any given time is involved exclusively in homosexuality.
Nonreproductive and Alternative Heterosexualities
As described above, male and female Guianan Cock-of-the-Rock have essentially no contact with each other outside of the mating season. Even during the breeding season, their interactions are often unfriendly or overtly aggressive. Males frequently harass females by chasing them around the display court and in some cases attempt to forcibly copulate with them. Females struggle violently during these rape attempts and are usually able to get away, but are visibly stressed following such interactions. In fact, only 20 percent of all lek visits by females result in copulation. There is also a significant proportion of nonbreeding individuals in the population: an average of 20 percent of adult males do not have display territories (and thus rarely, if ever, court or mate with females), while nearly two-thirds of territorial males fail to mate each year. Moreover, males are rarely able to acquire their own territories (and therefore court females) before they are three to four years old. In addition, many females who visit the courtship grounds never actually mate with males during the entire breeding season.
{569}
Sources
(* asterisked references discuss homosexuality/transgender)
Endler, J. A., and M. Thery (1996) "Interacting Effects of Lek Placement, Display Behavior, Ambient Light, and Color Patterns in Three Neotropical Forest-Dwelling Birds." American Naturalist 148:421-52.
Gilliard, E. T. (1962) "On the Breeding Behavior of the Cock-of-the-Rock (Aves, Rupicola rupicola)." Bulletin of the American Museum of Natural History 124:31-68.
Trail, P. W. (1989) "Active Mate Choice at Cock-of-the-Rock Leks: Tactics of Sampling and Comparison." Behavioral Ecology and Sociobiology 25:283-92.
* --- (1985a) "A Lek's Icon: The Courtship Display of a Guianan Cock-of-the-Rock." American Birds 39:235-40.
--- (1985b) "Courtship Disruption Modifies Mate Choice in a Lek-Breeding Bird." Science 227:778-80.
* --- (1983) "Cock-of-the-Rock: Jungle Dandy." National Geographic Magazine 164:831-39.
* Trail, P. W., and D. L. Koutnik (1986) "Courtship Disruption at the Lek in the Guianan Cock-of-the-Rock." Ethology 73:197-218.
 Cotingas
Cotingas
CALFBIRD (Perissocephalus tricolor)
IDENTIFIfATION: A crow-sized bird with cinnamon-brown plumage and a bare, blue-gray face. DISTRIBUTION: North-central South America, including Venezuela, the Guianas, Amazonian Brazil. HABITAT: Tropical forest. STUDY AREAS: Brownsberg Nature Park, Suriname; near the Kanuku Mountains, Guyana.
Social Organization
Calfbird males congregate, eight to ten at a time, on a display area or LEK in the understory of the rain forest canopy. Each lek consists of a central perch occupied by only one male, around which the remaining males cluster. Females (and males) visit the central male during courtship, usually at dawn; mating is polygamous or promiscuous, and females nest and raise the young on their own.
Description
Behavioral Expression: Male Calfbirds sometimes court other males who are attracted to the leks. A homosexual courtship begins when one male approaches {570}
the display site in a posture that females also adopt when entering the lek: the body is held in a horizontal position and all of the feathers are sleeked down, including the cowl feathers. He is attracted by the dramatic display of the central male, which consists of a loud, droning MOO call (hence the bird's name) sounding something like grr-aaa-oooo. While mooing, the displaying male first puffs himself up, fluffing up his cowl feathers, and then sinks back down in a bowing motion while exposing two bright orange feather globes on either side of his tail. The male in horizontal posture gets close to the mooing bird, who may then direct his courtship displays directly at the other male — for example, by hopping closer while mooing intensely, or engaging in ritualized wing-preening. Usually the approaching male is then chased away if he gets too close, and in fact no homosexual copulations have yet been observed in this species. However, heterosexual mating itself is fairly rare (only occurring in 14 percent of dawn courtship visits by females), and displaying males also frequently chase females who approach them. Furthermore, males do sometimes mount or attempt to mount other males outside of the lek, though this may occur in an aggressive context. Calfbirds also typically form "companionships" with each other: the two males perform coordinated courtship displays, almost touching as they perch side by side, while they both moo and bow in precise alternation. In some cases, companions never perform courtship displays without their male partner. Companions also travel together on the lek and may even share a "home" with each other. Calfbirds typically have what is known as a RETREAT, a special location or tree where each male regularly spends time when not on the lek. Display partners sometimes share the same retreat.
Female Calfbirds also develop bonds with one another: the two companions keep each other company while feeding, travel together to and from the lek, and even nest close to each other. This is all the more remarkable considering that there are no heterosexual pair-bonds in this species. Female pairs use a number of distinctive calls to communicate with each other. While feeding together, for example, they maintain contact with soft wark calls. When one female sits on her nest, the other may perch nearby uttering a rasping waaa call regularly for over an hour, perhaps acting as a lookout for her companion. And two females nesting close to each other sometimes communicate with a unique low, growling call that sounds like grewer grewer, which is only heard in this context. Although no same-sex courtship or copulation has yet been observed, females do sometimes perform behaviors that are typically associated with displaying males. For example, one female was seen to call and posture repeatedly in the fashion of a male courting on a lek (though she did this away from the lek, at her nest). Her call was like the first half of the male's mooing, and she displayed the orange tail ornaments usually seen in the male's courtship sequence. Her voice was also raspier than other females', and she built an exceptionally large nest. In addition, females sometimes adopt the male's characteristic upright and fluffed appearance when they visit the lek.
Frequency: Approximately 1-2 percent of all courtship visits involve one male displaying to another, and all males (except for the central one) have male display {571}
partners. Mounting attempts between males occur fairly frequently, but it is not known how prevalent female same-sex activities are.
Orientation: Because only the central male ever copulates with females, the remainder of the male population is effectively involved only in same-sex activities for the duration of the breeding season, whether display companionships or courtship/mounting with other males. The central male, however, displays mostly to females and occasionally to males, so his behavior is bisexual — albeit primarily heterosexual. In addition, in the next breeding season another male may become the central one, so at least some of the males exhibit sequential bisexuality, while the remainder may continue to experience longer periods of more exclusive homosexuality.
Nonreproductive and Alternative Heterosexualities
As discussed above, the majority of the male population in any given year does not reproduce: only one male (the central one in the lek) ever breeds with a female. Moreover, only 12 percent of females mate during their dawn courtship visits, and heterosexual relations are frequently fraught with difficulty. Copulations are brief, often incomplete, and accompanied by aggression. Females visiting the lek are constantly chased by the noncentral males, while nearly one-third of male-female copulations and more than half of all courtship visits are disrupted and harassed by other Calfbirds.
Sources
(* asterisked references discuss homosexuality/transgender)
* Snow, B. K. (1972) "A Field Study of the Calfbird Perissocephalus tricolor." Ibis 114:139-62.
--- (1961) "Notes on the Behavior of Three Cotingidae." Auk 78:150-61.
Snow, D. (1982) The Cotingas: Bellbirds, Umbrellabirds, and Other Species. Ithaca: Cornell University Press.
* --- (1976) The Web of Adaptation: Bird Studies in the American Tropics. New York: Quadrangle/New York Times Book Co.
* Trail, P. W. (1990) "Why Should Lek-Breeders Be Monomorphic?" Evolution 44:1837-52.
{572}
 Manakins
Manakins
SWALLOW-TAILED MANAKIN (Chiroxiphia caudata)
IDENTIFICATION: Adult males are bright blue with an orangish red crown and a black head and wings; females and yearling males are all green, while younger adult males are green or blue-green with an orange crown. DISTRIBUTION: Eastern Brazil, Paraguay, northwestern Argentina. HABITAT: Moist forests. STUDY AREA: Southeastern Brazil.
BLUE-BACKED MANAKIN (Chiroxiphia pareola)
IDENTIFICATION: Adult males are black with a red crown and a light blue patch on the back; yearling males and females are all green, while younger adult males are green with a reddish crown. DISTRIBUTION: Tobago, the Guianas, the Amazon Basin. HABITAT: Forest undergrowth, woodland. STUDY AREA: Tobago, West Indies; subspecies
C.p. atlantica.
Social Organization
Male Swallow-tailed Manakins form stable, long-term associations of four to six individuals that spend virtually all of their time together; they remain with each other throughout the mating season and usually from year to year as well. These groups display together on traditional courts or LEKS. The mating system is polygamous, with males (and probably also females) copulating with multiple partners, although large numbers of males are nonbreeders. Blue-backed Manakins have a similar social organization.

Homosexual courtship in a group of Swallow-tailed Manakins: one male (right) hovers in front of a younger male as the other males prepare to perform their part in the "jump display"
Description
Behavioral Expression: Male Swallow-tailed Manakins perform an elaborate group courtship ritual, the JUMP DISPLAY. Sometimes this display is directed toward females, sometimes toward yearling or younger adult males (the latter resembling females in their green plumage, but distinct because of their red-orange caps), and less commonly toward another adult male. To begin a homosexual jump display, a group of two to three adult males gathers on a display perch on their lek, lined up side by side, with the (younger) male they are displaying to perched at the head of this row. While uttering squawking notes that resemble a chorus of frogs, the adult {573}
males crouch down, quiver their bodies, and shuffle their feet, forming a collective vibrating mass. The courting males then take turns jumping up and hovering in front of the younger male while giving a sharp dik-dik-dik call, landing next to him. As each male jumps, the others slide down the perch toward his former position, repeating the sequence each time to produce a continuous rotation of flying and sliding birds, whose orange-red crowns collectively form a "whirling torch" in front of the courted bird. During this coordinated display, the younger male usually sits motionless in an upright posture. Males also sometimes mount each other and attempt copulation.
A similar revolving courtship display is performed by pairs of male Blue-backed Manakins for a third bird, which is sometimes a younger male. The display begins with a duet between the displaying males in order to attract a potential mate: the two birds perch side by side and utter perfectly synchronized chup calls. Once a younger male arrives, they begin courting him with their CATHERINE WHEEL DISPLAY. Each adult male jumps up alternately and flutters backward, "leapfrogging" over his partner, who moves forward to replace him in a precisely timed sequence that resembles two juggling balls. This is repeated up to 60 times to create a "cartwheel" of flying birds, oriented toward the male being courted, that gradually gets faster and faster. All the while the displaying males utter vibrant nasal or buzzing sounds that resemble the twanging of a Jew's harp. At the peak of the dancing frenzy, one of the adult males calls sharply and his partner disappears; the first male then begins courting the younger male one-on-one. He crisscrosses his display perch in a buoyant, butterfly-like flight, alighting in front of the other male every once in a while to lower his head, vibrate his wings and opalescent blue back, and display his brilliant red crest with its two extendable horns. During this performance, the younger male crouches and constantly turns to face the older male courting him, sometimes also responding with a similar bouncing "butterfly" flight across the display perch.
{574}
Frequency: The overall proportion of courtships or mounts that occur between males is not yet known for either of these species, although they may be relatively infrequent. However, two of three observed "butterfly" courtship displays in a study of Blue-backed Manakins were directed toward males, while in a two-year study of Swallow-tailed Manakins, only ten heterosexual matings were recorded. It is possible, therefore, that same-sex activity represents a sizable proportion of all courtship and/or sexual activity.
Orientation: Only one out of every four to six male Swallow-tailed Manakins displaying on the lek ever copulates with females; thus, a large proportion of males are not involved directly in male-female sexual (mounting) activity, although they do participate in both heterosexual and homosexual courtships. A similar pattern occurs in Blue-backed Manakins.
Nonreproductive and Alternative Heterosexualities
In addition to significant numbers of nonbreeding males, Swallow-tailed Manakin populations are characterized by several other less-than-optimal heterosexualities. Only about a third of male-female courtships ever result in mating: half of the time, females being courted simply leave, while in the remainder of cases other males disrupt the courtship displays. However, when mating does take place, a female may copulate with the same male up to six times in one visit; the same is also true for Blue-backed Manakins.
Sources
(* asterisked references discuss homosexuality/transgender)
Foster, M. S. (1987) "Delayed Maturation, Neoteny, and Social System Differences in Two Manakins of the Genus Chiroxiphia." Evolution 41:547-58.
--- (1984) "Jewel Bird Jamboree." Natural History 93(7):54-59.
--- (1981) "Cooperative Behavior and Social Organization in the Swallow-tailed Manakin (Chiroxiphia caudata)." Behavioral Ecology and Sociobiology 9:167-77.
Gilliard, E. T. (1959) "Notes on the Courtship Behavior of the Blue-backed Manakin (Chiroxiphia pareola )." American Museum Novitates 1942:1-19.
* Sick, H. (1967) "Courtship Behavior in the Manakins (Pipridae): A Review." Living Bird 6:5-22.
* --- (1959) "Die Balz der Schmuckvogel (Pipridae) [The Mating Ritual of Jewel Birds]." Journal fur Ornithologie 100:269-302.
* Snow, D. W. (1976) The Web of Adaptation: Bird Studies in the American Tropics. New York: Quadrangle /New York Times Book Co.
--- (1971) "Social Organization of the Blue-backed Manakin." Wilson Bulletin 83:3-38.
* --- (1963) "The Display of the Blue-backed Manakin, Chiroxiphia pareola, in Tobago, W.I." Zoologica 48:167-76.
{575}
 Antbirds
Antbirds
BICOLORED ANTBIRD (Gymnopithys bicolor)
OCELLATED ANTBIRD (Phaenostictus mcleannani)
IDENTIFICATION: Small (5-7 inch) birds with brown and rufous plumage and a bluish gray patch around the eyes; Ocellateds have a distinctive scalloped pattern on the back feathers. DISTRIBUTION: Central America and northwestern South America from Honduras to Ecuador. HABITAT: Rain forest undergrowth. STUDY AREA: Barro Colorado Island, Panama.
Social Organization
Antbirds get their name because they follow large swarms of army ants for food, often in flocks containing several different bird species. Both Bicolored and Ocellated Antbirds form monogamous pairs that are generally long-lasting. In addition, Ocellated Antbirds live in a complex extended family or "clan" structure, typically containing up to three generations of males and their mates. Females emigrate from these units to join other clans, while males often return to their extended family once they have found mates.
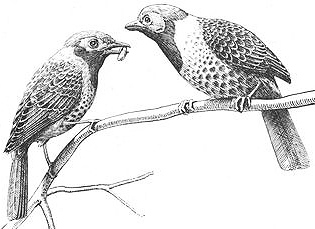
A male Ocellated Antbird (left) "courtship-feeding" his male pair-mate
Description
Behavioral Expression: Male homosexual pairs are a distinctive feature of Bicolored and Ocellated Antbird social life. Usually the pair-bond is initiated and strengthened (as in heterosexual pairs) when one male COURTSHIP-FEEDS the other, that is, ritually presents him with a "gift" of food (usually an insect or spider). In Ocellated Antbirds, males who court each other this way often adopt a characteristic pose (also used in heterosexual courtship) — ruffled throat feathers, stiff neck and upright posture, fluffed-out body feathers, with tail and legs spread. They also CAROL, producing a series of up to 15 whistled notes that decrease in pitch (sounding like chee chee chew chew). Unlike heterosexual mates, male partners typically reciprocate courtship feeding by passing the food gift back and forth between them. Homosexual courtship-feeding in Bicolored Antbirds is distinct from the heterosexual pattern in a number of other ways as well: either partner may initiate the activity in a male pair, and courtship-feeding is usually accompanied by CHIRPING — brief, musical cheup notes. In heterosexual pairs, only the male {576}
feeds the female, and the partners typically utter GROWLS during courtship-feeding (a rapid hissing or grunting noise composed of rough chauhh notes). Once paired, two males are constant companions, visiting ant swarms together and foraging side by side much as opposite-sex mated pairs do. Homosexual pair-bonds are sometimes long-lasting associations, persisting for many years. Partners may both be adults, or an older bird may pair with a younger one. In Ocellated Antbirds, father-son courtship-feedings also sometimes occur when a younger male remains within the clan structure.
Frequency: In Bicolored Antbirds, an estimated 2-3 percent of all pairs in some populations are composed of two males, and homosexual pairs may constitute up to 4-6 percent of the total number of pairs in any given year. The incidence of male pairs in Ocellated Antbirds is probably comparable.
Orientation: Approximately 5-14 percent of male Bicolored Antbirds may participate in a homosexual pairing or courtship at some point in their life. In both species, some males exhibit sequential bisexuality. In a few cases, for example, males have been mated to a female prior to their homosexual pairing (they may even have fathered young, and some are widowers), or they may go on to mate with a female when a homosexual pairing breaks up. Other males, however, show no signs of participating in heterosexual mating, and these birds may be involved exclusively in homosexual pairs, at least for a portion of their lives.
Nonreproductive and Alternative Heterosexualities
In both Bicolored and Ocellated Antbirds, significant numbers of birds are nonbreeding. As many as 45 percent of adult Bicolored males may not be heterosexually paired in any given year, and some males fail to acquire a mate for six or more years in a row. Younger males may delay their reproductive careers for up to a year after reaching sexual maturity — by remaining "at home" in their clans (Ocellateds) or wandering solitarily (Bicoloreds). In addition, male Antbirds have been known to live to a relatively old age — more than 11 years in Bicoloreds and Ocel-lateds. For a few individuals who have lost their female partners (either through death or divorce), this may be a postreproductive period in their lives. In addition to females leaving their mates for younger males, divorce and mate-switching also occasionally happen in adult Bicolored and newly paired Ocellated Antbirds. Often, a divorce is {577}
initiated by "extramarital" courtship between a mated female and an unpaired male. The extended families of Ocellated Antbirds also sometimes break up: heterosexual pairs wander off from the clan if they have not been able to breed successfully, or grandparents isolate themselves from their relatives. Heterosexual relations within a pair are not always smooth either: in Bicoloreds, for example, males are often distinctly hostile to their female mates. This is especially true early in their pair-bond, when he may aggressively "blast" her off her perch with hissing and snapping. Female Ocellated Antbirds have also been observed steadfastly refusing the courtship and copulatory advances of males. Finally, a number of incestuous activities have been documented in these species, including courtship and attempted copulation between Ocellated males and their mothers, and courtship of Bicolored daughters by their fathers.
Sources
(* asterisked references discuss homosexuality/transgender)
Willis, E.O. (1983) "Longevities of Some Panamanian Forest Birds, with Note of Low Survivorship in Old Spotted Antbirds (Hylophylax naevioides)." Journal of Field Ornithology54:413-14.
* --- (1973) The Behavior of Ocellated Antbirds. Smithsonian Contributions to Zoology no. 144. Washington, D.C.: Smithsonian Institution Press.
* --- (1972) The Behavior of Spotted Antbirds. Ornithological Monographs 10. Washington, D.C.: American Ornithologists' Union.
* --- (1967) The Behavior of Bicolored Antbirds. University of California Publications in Zoology 79. Berkeley: University of California Press.
 Tyrant Flycatchers
Tyrant Flycatchers
OCHER-BELLIED FLYCATCHER (Mionectes oleagineus)
IDENTIFICATION: A small (5 inch), plain olive-green bird with a long tail and an ocher- or tawny-colored lower breast. DISTRIBUTION: From Mexico south to the Amazon in South America; Trinidad and Tobago. HABITAT: Humid lowland forests, open shrubbery. STUDY AREA: Corcovado National Park, Costa Rica; subspecies
M.o. dyscola.
{578}
Social Organization
Ocher-bellied Flycatchers have a complex social organization, with three distinct categories of males. About 42 percent of males are TERRITORIAL, defending "courts" in the foliage within which they perform courtship displays; sometimes groups of two to six territorial males display close to each other in a LEK formation. Another 10 percent of males are SATELLITES, who associate with territorial males but do not display; they often eventually inherit the territory themselves. Finally, 48 percent of males are FLOATERS, who travel widely and do not hold any territories themselves. The mating system of Ocher-bellies is polygamous or promiscuous. No male-female pair-bonds are formed; instead, males mate with as many females as they can, but the female raises the young on her own.
Description
Behavioral Expression: Female Ocher-bellied Flycatchers are usually attracted to males displaying and singing on their territories, but sometimes another male approaches and is courted by the territorial male. The approaching male behaves much like a female, moving toward the center of the display court flicking his wings while the other male sings more intensely (making whistlelike notes), crouching and wing-flicking. The territorial male then trails the other male, following him closely and sometimes making soft ipp calls. The courtship sequence continues as in a heterosexual encounter with a series of three types of flight displays by the territorial male. The HOP DISPLAY involves the male bouncing excitedly back and forth between two perches uttering an eek call. In the FLUTTER FLIGHT, the displaying male traverses an arc between two perches with a special, slow wing-fluttering pattern, while in the HOVER FLIGHT, the male slowly rises in a hover above his perch or between two perches, often fairly close to the other male. The courtship sequence typically ends abruptly with the territorial male chasing the other male off while making chur calls.
Frequency: Approximately 17 percent of courtship sequences involve a male displaying to another male, and about 5 percent of male visits to territories result in courtship. Although no mountings or attempted copulations between males have been seen, heterosexual matings have rarely been witnessed either. At one study site, for example, only two male-female mountings were observed during more than 560 hours of observation over ten months.
Orientation: It is difficult to determine the relative proportion and "preference" of heterosexual versus homosexual behavior in Ocher-bellied Flycatchers. Some researchers believe that territorial males who court other males do not realize they are displaying to a bird of the same sex, in which case they would be exhibiting superficially heterosexual behavior toward (behaviorally) "transvestite" birds. For the males who approach territorial males, however, the situation is even less clear: many of these are probably floaters who presumably are aware that they are being courted by another male, i.e., they are ostensibly participating in {579}
homosexual activity. However, in at least one case the approaching male was a neighboring territorial male who also displayed to females on his own territory, i.e., his courtship interactions were actually bisexual.
Nonreproductive and Alternative Heterosexualities
As noted above, more than half of the male population consists of nonbreeders, since floaters and satellites rarely, if ever, mate heterosexually. Moreover, the absence of breeding activity in these males cannot be attributed to a shortage of available display sites, since more than three-quarters of suitable territories go unused (and nearly a quarter of these are especially prime pieces of "real estate").
Sources
(* asterisked references discuss homosexuality/transgender)
Sherry, T. W. (1983) "Mionectes oleaginea." In D. H. Janzen, ed., Costa Rican Natural History, pp. 586-87. Chicago: University of Chicago Press.
Skutch, A. E. (1960) "Oleaginous pipromorpha." In Life Histories of Central American Birds II, Pacific Coast Avifauna no. 34, pp. 561-70. Berkeley: Cooper Ornithological Society.
Snow, B. K., and D. W. Snow (1979) "The Ocher-bellied Flycatcher and the Evolution of Lek Behavior." Condor 81:286-92.
Westcott, D. A. (1997) "Neighbors, Strangers, and Male-Male Aggression as a Determinant of Lek Size." Behavioral Ecology and Sociobiology 40:235-42.
--- (1993) "Habitat Characteristics of Lek Sites and Their Availability for the Ocher-bellied Flycatcher, Mionectes oleagineus." Biotropica 25:444-51.
--- (1992) "Inter- and Intra-Sexual Selection: The Role of Song in a Lek Mating System." Animal Behavior 44:695-703.
* Westcott, D. A., and J. N. M. Smith (1994) "Behavior and Social Organization During the Breeding Season in Mionectes oleagineus, a Lekking Flycatcher." Condor 96:672-83.
{580}
SWALLOWS, WARBLERS, FINCHES, AND OTHERS
 Swallows
Swallows
TREE SWALLOW (Tachycineta bicolor)
IDENTIFICATION: A small to medium-sized swallow with iridescent blue-green upperparts, white underparts, and a tail that is only slightly forked. DISTRIBUTION: Canada and northern United States; winters in southern United States to northwestern South America. HABITAT: Forests, fields, meadows, marshes; usually near water. STUDY AREA: Allendale, Michigan.
Social Organization
Tree Swallows are extremely social birds: outside of the mating season, they gather in large flocks that may contain several hundred thousand individuals, while during the breeding season they form pairs and often nest in aggregations or colonies.
Description
Behavioral Expression: Groups of male Tree Swallows sometimes pursue other males during the mating season in order to copulate with them. When the object of their attentions alights, the males hover in a "cloud" above him, constantly fluttering and making the distinctive tick-tick-tick call that is characteristic of males when they are mating with females. If one of them succeeds in mounting the male, a complete homosexual copulation ensues: the male on top holds on to the other male's neck and back feathers with his bill, while the male being mounted lifts his tail so that genital contact can occur. As in heterosexual mating, multiple, repeated genital contacts may occur during a single copulation between two males, which can last for up to a minute (male-female mounts generally last about 30 seconds). The cluster {581}
of males may also engage in several consecutive episodes of homosexual mating: when the male who was mounted flies off, the group will continue pursuing him until he lands again, and the whole process is repeated.
Frequency: Homosexual copulations have been observed only occasionally in Tree Swallows. However, heterosexual nonmonogamous matings are also rarely seen in this species, yet they are known to be very common because of the high rate of offspring that result from them (see below). Most such copulations therefore probably occur in locations where (or at times when) they are not readily observed. Homosexual matings (which follow the pattern of nonmonogamous copulations) probably also occur more often than they are observed.
Orientation: Some males that participate in homosexual pursuits and copulations are probably bisexual: for example, one male who was mounted by other males was the father of six-day-old nestlings when he participated in homosexual activity. The males mounting him were not paired with female mates in the same nesting colony, however, and may have been nonbreeders (although they could also have been heterosexually paired birds visiting from another colony).
Nonreproductive and Alternative Heterosexualities
Heterosexual pairs of Tree Swallows sometimes copulate well before the female is fertile, and nonreproductive matings may also occur after the eggs are laid or even following hatching of chicks. Overall, each pair copulates about 50-70 times per clutch of eggs produced. At least 15 percent of matings also occur after the female's fertile period, and more than 20 percent of mounts do not involve genital contact. In addition, a large number of heterosexual copulations that take place during incubation — as well as throughout the breeding season — are nonmonogamous matings between a female and a male other than her mate. Although many pairs are monogamous in this species (about half of all females are strictly faithful), promiscuous copulations are a prominent feature of Tree Swallow heterosexual interactions. Females often solicit such copulations (sometimes from several different males) and are also able to effectively terminate unwanted promiscuous matings. They do this by flying away, refusing to lift their tail for genital contact, or turning their head and snapping or "chattering" at the male. Nonmonogamous matings frequently result in offspring: in some populations, 50-90 percent of all nests contain young that are not genetically related to their mother's mate, and these constitute 40-75 percent of all nestlings. In some families, all the offspring are fathered by other males. The opposite situation also sometimes occurs: youngsters may be related to the father but not his female partner. This may result from mate-switching (divorce and remating), or because females occasionally lay eggs in another female's nests (5-9 percent of all nests are PARASITIZED this way).
In some populations, 3-8 percent of males form polygamous trios in which they bond and breed with two females simultaneously. If the two females share a nest, one may help care for the other's nestlings if her own eggs do not hatch. Many {582}
populations also have large numbers of nonbreeding birds, sometimes called FLOATERS because they do not occupy their own territories and tend to travel widely. As many as a quarter of all reproductively mature females are floaters. In addition to helping raise unrelated birds of their own species, Tree Swallows sometimes "adopt" nests belonging to other birds such as purple martins (Progne subis) or bluebirds (Sialia spp.), raising the foster young in addition to their own. More than half of all Tree Swallow heterosexual pairs do not remain together for more than one breeding season. Single parenting also occasionally occurs in this species, for example if one parent is killed or dies during the breeding season. Frequently, however, the widowed parent re-pairs with another mate. If a single parent is laying or incubating eggs, its new mate often adopts the brood, but if a single parent already has nestlings from the previous mate, the new partner often kills them (usually by pecking) in order to begin breeding himself or herself. Infanticide also sometimes occurs when a female kills a paired female's nestlings in order to try to precipitate a divorce and mate with her partner.
Sources
(* asterisked references discuss homosexuality/transgender)
Barber, C. A., R. J. Robertson, and P. T. Boag (1996) "The High Frequency of Extra-Pair Paternity in Tree Swallows Is Not an Artifact of Nestboxes." Behavioral Ecology and Sociobiology 38:425-30.
Chek, A. A., and R. J. Robertson (1991) "Infanticide in Female Tree Swallows: A Role for Sexual Selection." Condor 93:454-57.
Dunn, P. O., and R. J. Robertson (1992) "Geographic Variation in the Importance of Male Parental Care and Mating Systems in Tree Swallows." Behavioral Ecology 3:291-99.
Dunn, P. O., and R. J. Robertson, D. Michaud-Freeman, and P. T. Boag (1994) "Extra-Pair Paternity in Tree Swallows: Why Do Females Mate with More than One Male?" Behavioral Ecology and Sociobiology 35:273-81.
Leffelaar, D., and R. J. Robertson (1985) "Nest Usurpation and Female Competition for Breeding Opportunities by Tree Swallows." Wilson Bulletin 97:221-24
--- (1984) "Do Male Tree Swallows Guard Their Mates?" Behavioral Ecology and Sociobiology 16:73-79.
Lifjeld, J. T., P. O. Dunn, R. J. Robertson, and P. T. Boag (1993) "Extra-Pair Paternity in Monogamous Tree Swallows." Animal Behavior 45:213-29.
Lifjeld, J. T., and R. J. Robertson (1992) "Female Control of Extra-Pair Fertilization in Tree Swallows." Behavioral Ecology and Sociobiology 31:89-96.
* Lombardo, M. P. (1996) Personal communication.
--- (1988) "Evidence of Intraspecific Brood Parasitism in the Tree Swallow." Wilson Bulletin 100:126-28.
--- (1986) "Extrapair Copulations in the Tree Swallow." Wilson Bulletin 98:150-52.
* Lombardo, M. P., R. M. Bosman, C. A. Faro, S. G. Houtteman, and T.S. Kluisza (1994) "Homosexual Copulations by Male Tree Swallows." Wilson Bulletin 106:555-57.
Morrill, S. B., and R. J. Robertson (1990) "Occurrence of Extra-Pair Copulation in the Tree Swallow (Tachycineta bicolor)." Behavioral Ecology and Sociobiology 26:291-96.
Quinney, T. E. (1983) "Tree Swallows Cross a Polygyny Threshold." Auk 100:750-54.
Rendell, W. B. (1992) "Peculiar Behavior of a Subadult Female Tree Swallow." Wilson Bulletin 104:756-59.
Robertson, R. J. (1990) "Tactics and Counter-Tactics of Sexually Selected Infanticide in Tree Swallows." In J. Blondel, A. Gosler, J.-D. Lebreton, and R. McCleery, eds., Population Biology of Passerine Birds: An Integrated Approach, pp. 381-90. Berlin: Springer-Verlag.
Robertson, R. J., B. J. Stutchbury, and R. R. Cohen (1992) "Tree Swallow (Tachycineta bicolor)." In A. Poole, P. Stettenheim, and F. Gill, eds., The Birds of North America: Life Histories for the 21st Century, no. 11. Philadelphia: Academy of Natural Sciences; Washington, D.C.: American Ornithologists' Union.
Stutchbury, B. J., and R. J. Robertson (1987a) "Signaling Subordinate and Female Status: Two Hypotheses for the Adaptive Significance of Subadult Plumage in Female Tree Swallows." Auk 104:717-23. {583}
--- (1987b) "Behavioral Tactics of Subadult Female Floaters in the Tree Swallow." Behavioral Ecology and Sociobiology 20:413-19.
--- (1987c) "Two Methods of Sexing Adult Tree Swallows Before They Begin Breeding." Journal of Field Ornithology 58:236-42.
--- (1985) "Floating Populations of Female Tree Swallows." Auk 102:651-54.
Venier, L. A., P. O. Dunn, J. T. Lifjeld, and R. J. Robertson (1993) "Behavioral Patterns of Extra-Pair Copulation in Tree Swallows." Animal Behavior 45:412-15.
Venier, L. A., and R. J. Robertson (1991) "Copulation Behavior of the Tree Swallow, Tachycineta bicolor: Paternity Assurance in the Presence of Sperm Competition." Animal Behavior 42:939-48.
 Swallows
Swallows
CLIFF SWALLOW (Hirundo pyrrhonota)
IDENTIFICATION: A bluish brown swallow with pale underparts, buff forehead, and a chestnut throat; tail is not forked. DISTRIBUTION: North and Central America; winters in southern South America. HABITAT: Open country, cliffs. STUDY AREAS: Near Jackson Hole (Moran), Wyoming, Lakeview, Kansas, and along the North and South Platte Rivers, Nebraska; subspecies H.p. hypopolia and H.p. pyrrhonota.
BANK SWALLOW (Riparia riparia)
IDENTIFICATION: A small, sparrow-sized swallow with a slightly forked tail, brown plumage, white underparts, and a brown breast band. DISTRIBUTION: Throughout North America and Eurasia; winters to South America and southern Africa. HABITAT: Open country near water. STUDY AREAS: Near Madison, Wisconsin, and Dunblane, Scotland; subspecies
R.r. riparia.
Social Organization
Cliff and Bank Swallows are highly gregarious and may flock by the hundreds or even thousands. They generally form mated pairs (although many alternative arrangements also occur, see below) and nest in colonies. Cliff Swallow colonies are the largest of any swallow in the world, often containing a thousand nests (and sometimes up to three times this number).
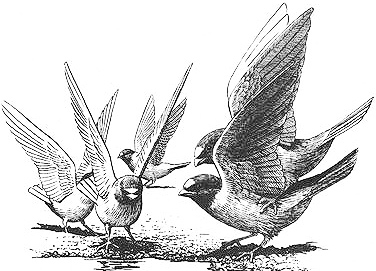
A male Cliff Swallow mounting another male (right) and attempting to copulate with him during a mud-gathering session
{584}
Description
Behavioral Expression: Male Cliff and Bank Swallows often try to copulate with both males and females that are not their own mates. Unlike in Tree Swallows, these are usually forced copulations or "rapes," since the bird being pursued — whether male or female — does not welcome the sexual advances of the male. Homosexual copulation attempts in Cliff Swallows take place on the ground at social gatherings of birds that are sunning themselves or gathering mud or grass for nests. Anywhere from a handful to several hundred individuals may be present at a time, although such groups usually contain 10-30 birds. At mud-gathering sessions, one male pounces on another male from above, landing on his back and often grabbing his head or neck feathers in his beak. At sunning sites, the male usually lands a few inches away from the other bird and makes threatening HEAD-FORWARD displays prior to jumping on his back. Once mounted, he spreads his tail and moves it sideways, trying to make cloacal (genital) contact, all the while vigorously flapping his wings; the other male usually strongly resists, and sometimes a fight ensues, before both birds fly off. The entire copulation attempt is usually quite brief, though it can last for up to 10 seconds (a relatively long duration for bird mountings). Forcible mountings of females follow this same pattern. When on the ground at mud-gathering sites, birds of both sexes typically flutter their wings above their backs to try to prevent males from mounting them.
Male Bank Swallows also pursue both females and males to try to copulate with them. Males first make many INVESTIGATORY CHASES of unfamiliar birds to determine if they are worth following. If they find a bird they are interested in — which may be another male — a full-fledged SEXUAL CHASE ensues, sometimes drawing several birds into the pursuit as well. Often the targeted bird is able to get away, but sometimes the chase ends with a forced copulation attempt. Homosexual mountings also occur when birds congregate on the ground, for instance to gather nest materials or dust themselves. At times, a veritable "orgy" develops as numerous males frantically try to mount birds of both sexes. Sometimes one or two males will even mount a third male who is already copulating with another bird.
{585}
Frequency: In Bank Swallows, 8 percent of sexual chases are homosexual, while 36-40 percent of investigatory chases involve males pursuing other males. Cliff Swallow rape attempts are extremely common, occurring every two to three minutes at some mud-gathering sites; one male may attempt to mount six to eight different birds in a five-to-ten-minute period. These mounts are probably fairly evenly distributed between males and females, although homosexual mounts may in fact occur more often. When presented with stuffed birds of both sexes in identical poses, male Cliff Swallows mounted other males nearly 65 percent of the time.
Orientation: Most, if not all, male Cliff and Bank Swallows that pursue copulations with other males also try to forcibly mount females, and to this extent they are bisexual. However, in Cliff Swallows only a few males appear to engage in such behavior with either males or females. Many of these are unpaired birds, although males who are already heterosexually paired (including fathers) also sometimes participate in promiscuous copulations.
Nonreproductive and Alternative Heterosexualities
Heterosexual social life in Cliff and Bank Swallows is replete with behaviors that deviate from the monogamous-pair/nuclear-family model. As discussed above, a large proportion of heterosexually mated Swallows seek copulations with birds other than their partner, and considerable evidence suggests that these mounting attempts are nonprocreative. Because of the resistance of the bird being mounted, the copulation is often not completed and sperm is rarely transferred. In addition, such attempts may also occur outside the breeding season when there is no possibility of fertilization. Genetic studies have shown that probably only 2-6 percent of Cliff Swallow nests have young that might result from nonmonogamous sexual activity. However, nearly a quarter of all nestlings, in more than half of all nests, are raised by birds other than their biological mother and/or father. This is because Cliff Swallows participate in an extraordinary array of activities that serve to exchange eggs and nestlings between families. For example, as many as 43 percent of all nests contain an egg laid by an outside female — this bird usually has her own nest, but she also PARASITIZES or adds eggs to other nests (and often has eggs added to her own as well). In some cases, this female may even return to the foreign nest to incubate the entire clutch (even though only one egg is hers), but she does not help raise the nestlings once they hatch. Often the intruding female's mate will destroy or toss out eggs in the host nest to make room for their own (up to 20 percent of all nests may suffer egg destruction). In other cases, males appear to destroy eggs in other nests so as to keep the laying female sexually receptive, thereby increasing the opportunities for heterosexual nonmonogamous matings. Birds also occasionally physically carry eggs from their own nest to another — about 6 percent of all nests contain eggs transferred this way. Finally, in a few cases Swallows have even been seen transferring actual baby birds between nests. Infanticide may also occur when birds attack and toss nestlings out of neighbors' nests. In both species, young birds gather into large
CRÈCHES or "day-care {586}
centers" — sometimes containing up to a thousand youngsters — which give them protection while their parents are away searching for food.
Divorce and single parenting also occur in Bank Swallows: females sometimes desert their mates to start a new family with another male, forcing their first mate to raise the nestlings on his own. In addition to the rape attempts described above, there is considerable aggression between the sexes as well. Ironically, male Bank Swallows often become violent toward their own female partners when trying to protect them from the advances of other males. Sometimes they knock their mate to the ground, fighting her directly, or try to force her back into their burrow. Nonreproductive sexual behaviors are also prevalent in these two species. Besides copulations outside the breeding season and group sexual activity (mentioned above), members of a Cliff Swallow pair often copulate at a rate far in excess of that needed simply to fertilize their eggs. In addition, males of both species have occasionally been seen trying to mate with dead birds, as well as with other species such as barn swallows (Hirundo rustica) and Tree Swallows.
Sources
(* asterisked references discuss homosexuality/transgender)
* Barlow, J. C., E. E. Klaas, and J. L. Lenz (1963) "Sunning of Bank Swallows and Cliff Swallows." Condor 65:438-48.
Beecher, M. D., and 1. M. Beecher (1979) "Sociobiology of Bank Swallows: Reproductive Strategy of the Male." Science 205:1282-85.
Beecher, M. D., I. M. Beecher, and S. Lumpkin (1981) "Parent-Offspring Recognition in Bank Swallows (Riparia riparia): I. Natural History." Animal Behavior 29:86-94.
Brewster, W. (1898) "Revival of the Sexual Passion in Birds in Autumn." Auk 15:194-95.
* Brown, C. R., and M.B. Brown (1996) Coloniality in the Cliff Swallow: The Effect of Group Size on Social Behavior. Chicago: University of Chicago Press.
* --- (1995) "Cliff Swallow (Hirundo pyrrhonota)." In A. Poole and F. Gill, eds., The Birds of North America: Life Histories for the 21st Century, no. 149. Philadelphia: Academy of Natural Sciences; Washington, D.C.: American Ornithologists' Union.
--- (1989) "Behavioral Dynamics of Intraspecific Brood Parasitism in Colonial Cliff Swallows." Animal Behavior 37:777-96.
--- (1988a) "A New Form of Reproductive Parasitism in Cliff Swallows." Nature 331:66-68.
--- (1988b) "The Costs and Benefits of Egg Destruction by Conspecifics in Colonial Cliff Swallows." Auk 105:737-48.
--- (1988c) "Genetic Evidence of Multiple Parentage in Broods of Cliff Swallows." Behavioral Ecology and Sociobiology 23:379-87.
Butler, R. W. (1982) "Wing-fluttering by Mud-gathering Cliff Swallows: Avoidance of 'Rape' Attempts?" Auk 99:758-61.
* Carr, D. (1968) "Behavior of Sand Martins on the Ground." British Birds 61:416-17.
Cowley, E. (1983) "Multi-Brooding and Mate Infidelity in the Sand Martin." Bird Study 30:1-7.
* Emlen, J. T., Jr. (1954) "Territory, Nest Building, and Pair Formation in Cliff Swallows." Auk 71:16-35.
--- (1952) "Social Behavior in Nesting Cliff Swallows." Condor 54:177-99.
Hoogland, J. L., and P. W. Sherman (1976) "Advantages and Disadvantages of Bank Swallow (Riparia riparia) Coloniality." Ecological Monographs 46:33-58.
* Jones, G. (1986) "Sexual Chases in Sand Martins (Riparia riparia): Cues for Males to Increase Their Reproductive Success." Behavioral Ecology and Sociobiology 19:179-85.
* Petersen, A. J. (1955) "The Breeding Cycle in the Bank Swallow." Wilson Bulletin 67:235-86.
Thom, A. S. (1947) "Display of Sand-Martin." British Birds 40:20-21.
{587}
 Wood Warblers
Wood Warblers
HOODED WARBLER (Wilsonia citrina)
IDENTIFICATION: A small songbird with bright yellow underparts, olive green upperparts, and a black crown ("hood") and throat in adult males and some females (see below). DISTRIBUTION: Eastern North America; winters in Mexico and Central America. HABITAT: Deciduous forests, cypress swamps. STUDY AREA: Smithsonian Environmental Research Center near Annapolis, Maryland.
Social Organization
During the breeding season, male Hooded Warblers establish and defend territories, attracting mates with whom they usually form pair-bonds. Outside of the mating season, birds migrate south to their winter homes, where the two sexes live largely separate from each other.

A pair of male Hooded Warblers tending their chicks
Description
Behavioral Expression: Male Hooded Warblers occasionally form homosexual pairs and become joint parents. Same-sex pair-bonds develop early in the mating season when one male is attracted to another male's territory by his singing. In some cases, this is a male he has previously "prospected" on a visit to his territory during the prior mating season. Once a pair-bond is established, the males focus their attentions on parenting duties. Homosexual couples acquire nests and eggs in a variety of ways. Some pairs may build their own nests: although male Hooded Warblers in heterosexual pairs rarely build nests, at least one bird in a homosexual pair was observed carrying grass fibers to a nest and shaping the cup by repeatedly sitting in the nest and shifting his position. It is not known, however, whether he had built the nest or was simply adding material to a nest built by another pair. As for eggs, some pairs incubate eggs laid by another species of bird, the Brown-headed Cowbird. This species is known as a PARASITE because it always lays its eggs in the nests of other birds, "forcing" them to raise its young. Hooded Warblers are particularly susceptible to parasitism by Cowbirds: in some populations, three-quarters of all nests are parasitized, and Cowbirds appear to prefer Hooded Warbler nests over those of other species. Cowbirds occasionally lay eggs in completely empty nests, so some homosexual pairs of Hooded Warblers may build their own {588}
nest and end up tending only Cowbird eggs. Usually, though, a Cowbird adds its egg(s) to a nest that already has eggs (often removing part of the original clutch). Sometimes a Hooded Warbler mother abandons her nest once it has been disturbed this way, and some homosexual pairs may "adopt" such abandoned nests, or else the father of such a nest may re-pair with a male following the mother's abandonment. At least two male pairs have been observed tending nests that were parasitized, since they each contained both Cowbird and Hooded Warbler chicks. Other male couples probably adopt nests that have been abandoned after attacks by predators. Bluejays and squirrels often prey on Hooded Warbler nestlings, and usually their mother will abandon the entire nest even if only one youngster has been taken. Finally, some homosexual pairs may tend eggs that have been directly laid in their nest by a female Hooded Warbler. In many bird species, females lay eggs in nests belonging to other birds of the same species (this is another form of parasitism); although this rarely occurs in Hooded Warblers, it is a possible source of eggs for homosexual pairs.
Once they have acquired a nestful of eggs, male couples typically divide up the parenting duties: one attends to nest repair, incubation of the eggs, and brooding of the nestlings, while the other feeds his mate and defends the territory. Both birds feed the nestlings insects such as crane flies. Although this division of labor is similar to that in heterosexual pairs — females typically build nests and incubate, males defend territories, and both feed nestlings — there are crucial differences. In homosexual pairs, incubating males are often fed by their mate, which occurs only rarely in heterosexual pairs. In addition, one male who engaged in typically "female" parental duties was later observed performing territorial singing (albeit with a song pattern that differed from that of most other males). Nests belonging to homosexual pairs are often lost entirely to predators, but up to 50 percent or more of all heterosexual nests are lost in the same way. The male couples that have been followed over a longer time do not appear to re-pair with the same mate in subsequent breeding seasons; their divorces may be related to the loss of nests to predators. Heterosexual divorce is also common in Hooded Warblers, with as many as half of all male-female pairs failing to remain together, perhaps also due in large part to loss of nests. It is possible as well that divorce is simply a general feature of {589}
pair-bonding in this species (heterosexual or homosexual) independent of nest losses, or that the particular pairs being studied happened to end in divorce without this being indicative of a larger pattern.
Some female Hooded Warblers are transvestite, having the same black hood that males do. In fact there is a continuum of transgendered physical appearance in females: some have no black feathers on their head at all, some have an intermediate amount with a black "bib" around the throat, while others are almost indistinguishable from males. In addition, a few females can sing (typically only males in this species are able to sing). Transgendered females usually mate with males and raise young just like nontransgendered females.
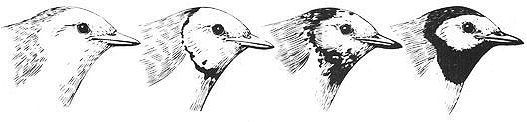
Transgendered Hooded Warblers: females of this species usually have little or no black on their heads (far left), but some individuals are plumage transvestites, exhibiting a full malelike black hood and chin strap (far right).
Other females exhibit a gradation of plumage patterns that fall between these two extremes (center).
Frequency: The overall incidence of homosexual pairs in Hooded Warblers is not known, since no widespread, systematic study has yet been conducted to determine their prevalence. However, in one population observed over three years, 4 percent of the pair-bonds (3 of 80) were between males. Although overt sexual behavior has not yet been observed between such pair-mates, heterosexual copulations (both within-pair and nonmongamous) are rarely seen in this species either; it is possible, therefore, that homosexual copulations do take place. Among females, plumage transvestism is a regular occurrence, as about 59 percent of females have some degree of malelike black feathers on their head: 40 percent have only a slight amount, 17 percent an intermediate amount, and 2 percent have a nearly complete black hood.
Orientation: Some male Hooded Warblers appear to be exclusively homosexual, pairing only with males; if they divorce a male partner, they re-pair with another male in subsequent breeding seasons. These males often perform parenting duties typically associated with females in heterosexual pairs. Other males, however, are bisexual, alternating between homosexual pairings and heterosexual ones in different breeding seasons.
Nonreproductive and Alternative Heterosexualities
As mentioned above, heterosexual divorce is common in Hooded Warblers, as are a number of other variations on the nuclear family and monogamous pair-bond. About 4 percent of males form trios, mating with two females who both nest simultaneously on the male's territory; 6 percent of males are nonbreeders, and some females remain single as well. Among paired birds, promiscuous copulations also occur very frequently: 30-50 percent of all females copulate with males other than their mates (usually neighboring males), and more than a third of all nestlings in some populations are fathered by a bird other than their mother's mate. In addition, males sometimes adopt young birds from neighboring families whose own parents have finished caring for them; adoptive fathers typically feed these youngsters along with their own nestlings. The adopted birds are usually not genetically related to their foster fathers, i.e., they are not the result of promiscuous matings by the bird who adopts them. Single parenting is also a regular occurrence in Hooded {590}
Warblers: once their young can fly, parents usually separate and each takes care of half the brood (unless the female begins a second family, in which case the male will assume responsibility for all of the youngsters). In fact, single parenting is generally more extensive and longer-lasting than male-female coparenting in this species. Nestlings receive biparental care for only eight or nine days, while single parenting can last for three to six weeks and involves feeding rates that are three to five times higher than that of coparents. As a result of separation of mated pairs, males and females are together for only about one month out of the entire year. During the winter, the two sexes occupy largely segregated habitats, with males preferring forests and females scrub areas.
Sources
(* asterisked references discuss homosexuality/transgender)
Evans Ogden, L. J., and B. J. Stutchbury (1997) "Fledgling Care and Male Parental Effort in the Hooded Warbler (Wilsonia citrina)." Canadian Journal of Zoology 75:576-81.
--- (1994) "Hooded Warbler (Wilsonia citrina):" In A. Poole and F. Gill, eds., The Birds of North America: Life Histories for the 21st Century, no. 110. Philadelphia: Academy of Natural Sciences; Washington, D.C.: American Ornithologists' Union.
Godard, R. (1993) "Tit for Tat Among Neighboring Hooded Warblers." Behavioral Ecology and Sociobiology 33:45-50.
--- (1986) "Long-Term Memory of Individual Neighbors in a Migratory Songbird." Nature 350:228-29.
* Lynch, J. F., E. S. Morton, and M. E. Van der Voort (1985) "Habitat Segregation Between the Sexes of Wintering Hooded Warblers (Wilsonia citrina)." Auk 102:714-21.
* Morton, E. S. (1989) "Female Hooded Warbler Plumage Does Not Become More Male-Like With Age." Wilson Bulletin 101:460-62.
* Niven, D. K. (1997) Personal communication.
* --- (1993) "Male-Male Nesting Behavior in Hooded Warblers." Wilson Bulletin 105:190-93.
Stutchbury, B. J. M. (1998) "Extra-Pair Mating Effort of Male Hooded Warblers, Wilsonia citrina." Animal Behavior 55:553-61.
* --- (1994) "Competition for Winter Territories in a Neotropical Migrant: The Role of Age, Sex, and Color." Auk 111:63-69.
Stutchbury, B. J., and L. J. Evans Ogden (1996) "Fledgling Adoption in Hooded Warblers (Wilsonia citrina): Does Extrapair Paternity Play a Role?" Auk 113:218-20.
* Stutchbury, B. J., and J. S. Howlett (1995) "Does Male-Like Coloration of Female Hooded Warblers Increase Nest Predation?" Condor 97:559-64.
Stutchbury, B. J., J. M. Rhymer, and E. S. Morton (1994) "Extrapair Paternity in Hooded Warblers." Behavioral Ecology 5:384-92.
{591}
 Finches
Finches
CHAFFINCH (Fringilla coelebs)
IDENTIFICATION: A sparrow-sized bird with olive-brown plumage, distinctive white shoulder bars, and (in males) blue-gray crown. DISTRIBUTION: Europe, Siberia, central Asia, North Africa. HABITAT: Forest, farmland. STUDY AREAS: Ylivieska, Finland; near Cambridge, England; subspecies F.c. coelebs and F.c. gengleri.
SCOTTISH CROSSBILL (Loxia scotica)
IDENTIFICATION: A sparrow-sized bird with olive to orange-red plumage and a distinctive crossed bill. DISTRIBUTION: Northern Scotland. HABITAT: Coniferous forest. STUDY AREAS: Speyside and Sutherland, Scotland.
Social Organization
Chaffinches and Scottish Crossbills commonly associate in flocks; the mating system involves (usually monogamous) pair-bonding.
Description
Behavioral Expression: Female Chaffinches sometimes form homosexual pair-bonds with each other. In this species, usually only males sing; however, in same-sex pairs, one female partner typically sings much in the manner of a male, throwing back her head while perched conspicuously in the trees (this is probably how she attracts her female mate). Her song resembles that of males in duration, loudness, and structure, consisting of a long stream of rapidly trilled notes of descending pitch, finished off with a staccato end phrase or "flourish." It differs from male song, however, in having not quite as ringing a tone. Like male Chaffinches, she may even COUNTERSING in response to a neighboring male's song, the two birds "replying" to each other with alternating or syncopated song phrases. Females in same-sex pairs may also try to sing together, although one partner may only be able to produce an incomplete version of the song. The two females behave like other mated couples, and may even engage in courtship pursuits known as SEXUAL CHASES. In this activity — which is a demonstration of sexual interest — one female zigzags after her partner using a special form of flying known as MOTH FLIGHT (rapid, shallow wingbeats without the pauses or undulating quality typical of regular flight). Occasionally, juvenile males pursue other males in such sexual chases. {592}
Homosexual couples also occur in Scottish Crossbills — but among males. A male attracts potential mates by singing high in a treetop, advertising his presence with a loud stream of notes transcribed as chip-chip-chip-gee-gee-gee chip-chip-chip. Most singing males respond aggressively to other males who enter their territory, but occasionally the displaying male does not chase away another male that is attracted to him. The two males may then pair up and associate in much the same way that heterosexual couples do, except that copulation has not been observed between male partners. They synchronize their movements, traveling together between forest clumps, sometimes one leading the other. Occasionally, two neighboring males who are each heterosexually mated become companions. The two forage together and jointly defend clumps of pine trees (their principal food source) while still maintaining their opposite-sex bonds. The two males may even visit their female mates together, attending first to one and then the other, with one male feeding his female partner while his male companion waits for him.
Frequency: In both of these species, homosexual pairs are probably only an occasional occurrence. Although no statistics concerning the incidence of same-sex pairing are available, in Chaffinches about 1 in 150 nests contains a SUPERNORMAL CLUTCH of 7-8 eggs. In many species such larger-than-average clutches are laid by female pairs, and this may also be true in Chaffinches (although their specific association with same-sex pairing has not yet been demonstrated).
Orientation: Individual Chaffinches or Scottish Crossbills who form same-sex associations have not been tracked throughout their entire lives to determine if they only pair with members of the same sex. However, since heterosexual pairs in these species are usually lifelong, it is not unlikely that homosexual pairs would be as well. In addition, some Scottish Crossbill males are bisexual, forming bonds with both males and females simultaneously.
Nonreproductive and Alternative Heterosexualities
A variety of nonprocreative heterosexual behaviors are found in Chaffinches and Scottish Crossbills. Chaffinch couples sometimes copulate during incubation (i.e., after fertilization has occurred), and only 40-50 percent of all matings during the fertilizable period involve full genital contact. Some pairs may attempt to mate more than 200 times for each clutch of eggs they produce, as often as 4 times an hour. Mounts are often not completed for a variety of reasons: the male may slip off the female's back or mount in a reversed head-to-tail position, or (more commonly) either partner may flee from the other out of fear or because of provocation from its mate. In fact, heterosexual courtship and mating in this species often entails a considerable amount of aggressive behavior between the sexes. Many birds do not participate in breeding at all: there are surplus flocks of single birds in many populations (males in Scottish Crossbills, both sexes in Chaffinches), as well as nonbreeding pairs in Scottish Crossbills. Interestingly, outside of the breeding season sexual segregation is also the rule. Males and female tend to migrate separately {593}
(wintering flocks are often same-sex), and females generally travel farther than males, often resulting in local populations with more males than females. In fact, this phenomenon has lent the Chaffinch its scientific name: coelebs means "bachelor" in Latin.
Scottish Crossbills frequently engage in "symbolic" matings early in the breeding season, in which the male mounts the female but no genital contact occurs. Sometimes these mounts are promiscuous: several males other than the female's mate may participate. Nonmonogamous matings are also fairly common in Chaffinches, often between neighboring birds. About 8 percent of all mating activity involves nonmates, and 17 percent of all offspring (in a quarter of all broods) result from such copulations. Occasionally, a pair will separate or "divorce" during the breeding season, after which the single female may copulate with several already paired males. Eventually, a polygamous trio may develop if one of these males forms a pair-bond with her in addition to his own mate. Some Scottish Crossbills are also polygamous, forming trios of one male with two females. In this species, parents (in monogamous pairs) regularly become single parents: they separate when their young are able to fly, dividing the young between them and raising them on their own. Because the mother and father often travel to widely separated feeding grounds, youngsters belonging to the same family may be raised in very different environments. Occasionally, unrelated birds — both males and females — help feed and "foster-parent" the young birds after their biological parents have separated.
Sources
(* asterisked references discuss homosexuality/transgender)
Adkisson, C. S. (1996) "Red Crossbill (Loxia curvirostra)." In A. Poole, P. Stettenheim, and F. Gill, eds., The Birds of North America: Life Histories for the 21st Century, no. 256. Philadelphia: Academy of Natural Sciences; Washington, D.C.: American Ornithologists' Union.
Halliday, H. (1948) "Song of Female Chaffinch." British Birds 41:343-44.
Hanski, I. K. (1994) "Timing of Copulations and Mate Guarding in the Chaffinch Fringilla coelebs." Ornis Fennica 71:17-25.
Kling, J. W., and J. Stevenson-Hinde (1977) "Development of Song and Reinforcing Effects of Song in Female Chaffinches." Animal Behavior 25:215-20.
Knox, A. G. (1990) "The Sympatric Breeding of Common and Scottish Crossbills Loxia curvirostra and L. scotica and the Evolution of Crossbills." Ibis 132:459-66.
* Marjakangas, A. (1981) "A Singing Chaffinch Fringilla coelebs in Female Plumage Paired with Another Female-Plumaged Chaffinch." Ornis Fennica 58:90-91.
* Marler, P. (1956) Behavior of the Chaffinch Fringilla coelebs. Behavior Supplement V. Leiden: E. J. Brill.
--- (1955) "Studies of Fighting in Chaffinches. 2. The Effect on Dominance Relations of Disguising Females as Males." British Journal of Animal Behavior 3:137-46.
* Nethersole-Thompson, D. (1975) Pine Crossbills: A Scottish Contribution. Berkhamsted: T. and A. D. Poyser.
Sheldon, B. C. (1994) "Sperm Competition in the Chaffinch: The Role of the Female." Animal Behavior 47:163-73.
Sheldon, B. C., and T. Burke (1994) "Copulation Behavior and Paternity in the Chaffinch." Behavioral Ecology and Sociobiology 34:149-56.
Svensson, B. V. (1978) "Clutch Dimensions and Aspects of the Breeding Strategy of the Chaffinch Fringilla coelebs in Northern Europe: A Study Based on Egg Collections." Ornis Scandinavica 9:66-83.
Voous, K. H. (1978) "The Scottish Crossbill Loxia scotica." British Birds 71:3-10.
{594}
 Shrikes, Tits, and Bluebirds
Shrikes, Tits, and Bluebirds
RED-BACKED SHRIKE (Lanius collurio)
IDENTIFICATION: A small (6 1/2 inch) bird with a thick, hooked bill, grayish brown plumage, and a darker facial mask (black in males). DISTRIBUTION: Throughout Eurasia and northeast Africa; winters to southern Africa. NABITAT: Savanna, woodland, scrub, farmland. STUDY AREA: New Forest, Hampshire, England.
BLUE TIT (Parus caeruleus)
IDENTIFICATION: A tiny (4 1/2 inch) chickadee-like bird with a bright blue crown, black-and-white face, bluish green plumage, and yellow underparts. DISTRIBUTION: Europe, Middle East, North Africa. HABITAT: Woodland, human habitation. STUDY AREA: Marley Wood, Oxfordshire, England; subspecies P.c. obscurus.
EASTERN BLUEBIRD (Sialia sialis)
IDENTIFICATION: A sparrow-sized bird with bright blue plumage, white underparts, and a chestnut throat and breast. DISTRIBUTION: North and Central America. HABITAT: Open woodland, orchards, farmland, pine savanna. STUDY AREA: In the town of Washington, Michigan.
Social Organization
Red-backed Shrikes generally establish monogamous pair-bonds during the mating season and occupy partially overlapping territories. Outside of the breeding season, the birds are more solitary, and males and females typically occupy segregated habitats (males in more open bush country, females in more dense woodland). Blue Tits and Eastern Bluebirds are also territorial and form pair-bonds, but may associate in flocks outside of the breeding season.

A nesting pair of female Red-backed Shrikes in England simultaneously incubating their eggs
Description
Behavioral Expression: In Red-backed Shrikes and Blue Tits, two females sometimes pair with each other, build a nest, and lay eggs. Both partners in Red-backed Shrike homosexual pairs take turns incubating the eggs (in heterosexual pairs, only the female incubates). Sometimes they even sit on the nest together, side by side or one partially covering the other. Both females also lay eggs: as a result, {595}
their nests have SUPERNORMAL CLUTCHES of 9-12 eggs, twice the number in most nests belonging to heterosexual pairs. If their nest is robbed by predators, the pair will dutifully build a second one (often in a much higher location, to make it inaccessible to most ground predators) and begin laying eggs again (as do male-female pairs when they lose a nest). However, all of the eggs they lay are typically infertile since neither female has mated with a male, and the pair usually abandons the nest after the eggs have failed to hatch. Blue Tit female pairs are unlike most homosexual pairs in birds because only one female lays eggs. As a result, nests belonging to such pairs contain not supernormal clutches, but exceptionally small clutches of 3 or so eggs (nests of male-female pairs typically contain about 11 eggs). Both females incubate the eggs simultaneously, sitting together on the nest facing in the same direction. As in Red-backed Shrike homosexual pairs, the eggs are usually infertile and the pair eventually abandons the nest.
In Eastern Bluebirds, two males sometimes associate with one another in what appears to be a homosexual pair-bond: they travel exclusively in each other's company (perhaps even spending the winter together), jointly inspect nest sites during the early spring, and court one another. The latter activity involves COURTSHIP-FEEDING — a behavior also seen in heterosexual pairs — in which one male offers a symbolic food gift, such as a cutworm, to the other male, often preceded by a distinctive call. Paired males might be related to each other (father-son or brothers). Occasionally, a female who has lost her heterosexual mate is joined by another female, who helps coparent her offspring. The two birds take turns feeding the youngsters and may also be assisted by one or more of their young from a previous brood.

Nests belonging to homosexual pairs of Black-winged Stilts (left) and Red-backed Shrikes (right). Both females in the pair lay eggs, and therefore their nests contain "supernormal clutches" (double the usual number of eggs). Because neither female has mated with males, however, these clutches typically consist entirely of unfertilized eggs.
Frequency: Same-sex pairs probably occur only sporadically in these three species: in Red-backed Shrikes, for example, female couples account for perhaps no more than 1 percent of all pairs.
Orientation: Female Red-backed Shrikes and Blue Tits are probably exclusively homosexual, at least for the time that they remain paired with their female partner, since they invariably lay eggs that are infertile (indicating no mating with males). Whether such birds ever subsequently form or have previously formed heterosexual pair-bonds is not known.
{596}
Nonreproductive and Alternative Heterosexualities
Approximately 17 percent of all sexual activity in opposite-sex pairs of Blue Tits occurs outside the female's fertile period. Male Eastern Bluebirds also sometimes try to copulate when females are nonfertile, such as during incubation or after the breeding season is over. Other nonprocreative matings involve occasional interspecies encounters: juvenile Eastern Bluebirds have been observed being mounted by great crested flycatchers (Myiarchus crinitus). Courtship and mounting between pair-bonded Eastern Bluebirds sometimes involve considerable aggression, with males attacking their partner and violently pecking her head or knocking her over with their feet. Heterosexual copulations are also occasionally interrupted by neighboring birds that attack the mating pair. Within-pair copulation rates are quite high in Red-backed Shrikes, roughly three times a day during the female's fertile period. In addition, nonmonogamous matings are common in these species: approximately 10 percent of Blue Tit copulations are promiscuous, involving birds that are not paired to each other. About a quarter of Bluebird nests and a third to nearly a half of all Blue Tit broods contain youngsters fathered by an outside bird. Promiscuous copulations also occasionally occur in Red-backed Shrikes, including forced matings or rapes in which males attack and mount females or violently push them off their perches. About 5 percent of Red-backed Shrike nestlings are fathered by a bird other than their mother's mate, compared to 8-35 percent in Eastern Bluebirds, and 11-14 percent in Blue Tits.
Several other alternative parenting arrangements occur in these species. Red-backed Shrike youngsters sometimes move to adjacent territories where they are adopted by other families, and a few nonbreeding birds of both sexes help feed chicks belonging to other pairs (sometimes even assuming full responsibility for them later on). "Helper" males such as these may even replace the biological father as a model for song-learning by the youngsters. In addition, late-breeding Red-backed Shrikes sometimes become single parents when their mate deserts them; occasionally two such birds and their offspring join together to form a "blended family." At least 15 percent of Eastern Bluebird mothers in some populations raise unrelated youngsters when outside females lay eggs in their nest. The divorce rate for Blue Tits varies considerably, from 8-85 percent (depending on the population); in addition, about a third of females and up to 20 percent of all males in some areas form polygamous trios (or occasionally quartets). Other more complex family arrangements also occur: one female Blue Tit was part of a polygamous trio with another female and a male; she also copulated promiscuously with a second male, eventually forming a new polygamous bond with both him and yet a third male, all of whom helped care for the young! Overall, however, more than 85 percent of Blue Tits never successfully breed, and about a third of all parents never have grandchildren. Red-backed Shrike parents occasionally cannibalize their own young or eggs.
Other Species
Some male Pied Flycatchers (Ficedula hypoleuca), especially younger ones, are transvestite, having the same brownish plumage that females do; these birds are sometimes courted by other males.
{597}
Sources
(* asterisked references discuss homosexuality/transgender)
Alsop, F. J., III (1971) "Great Crested Flycatcher Observed Copulating with an Immature Eastern Bluebird." Wilson Bulletin 83:312.
* Ashby, E. (1958) "Incidents of Bird Life [report on Red-backed Shrikes]." The Countryman: A Quarterly Review and Miscellany of Rural Life and Progress 55:272.
Birkhead, T. R., and A. P. Moller (1992) Sperm Competition in Birds: Evolutionary Causes and Consequences. London: Academic Press.
* Blakey, J. K. (1996) "Nest-Sharing by Female Blue Tits." British Birds 89:279-80.
* Cramp, S., and C. M. Perrins, eds. (1993) "Red-backed Shrike (Lanius collurio)." In Handbook of the Birds of Europe, the Middle East, and North Africa, vol. 7, pp. 456-78. Oxford: Oxford University Press.
Dhondt, A. A. (1989) "Blue Tit." In I. Newton, ed., Lifetime Reproduction in Birds, pp. 15-33. London: Academic Press.
--- (1987) "Reproduction and Survival of Polygynous and Monogamous Blue Tit Parus caeruleus." Ibis 129:327-34.
Dhondt, A. A., and F. Adriaensen (1994) "Causes and Effects of Divorce in the Blue Tit Parus caeruleus." Journal of Animal Ecology 63:979-987.
Fornasari, L., L. Bottoni, N. Sacchi, and R. Massa (1994) "Home-Range Overlapping and Socio-Sexual Relationships in the Red-backed Shrike Lanius collurio." Ethology, Ecology, and Evolution 6:169-177.
Gowaty, P. A., and W. C. Bridges (1991) "Behavioral, Demographic, and Environmental Correlates of Extrapair Fertilizations in Eastern Bluebirds, Sialia sialis." Behavioral Ecology 2:339-50.
Gowaty, P. A., and A. A. Karlin (1984) "Multiple Maternity and Paternity in Single Broods of Apparently Monogamous Eastern Bluebirds (Sialia sialis)." Behavioral Ecology and Sociobiology 15:91-95.
Hartshorne, J. M. (1962) "Behavior of the Eastern Bluebird at the Nest." Living Bird 1:131-49.
Herremans, M. (1997) "Habitat Segregation of Male and Female Red-backed Shrikes Lanius collurio and Lesser Gray Shrikes Lanius minor in the Kalahari Basin, Botswana." Journal of Avian Biology 28:240-48.
Jakober, H., and W. Stauber (1994) "Kopulationen und Partnerbewachung beim Neuntoter Lanius collurio [Copulation and Mate-Guarding in the Red-backed Shrike]." Journal fur Ornithologie 135:535-47.
--- (1983) "Zur Phanologie einer Population des Neuntoters (Lanius collurio) [On the Phenology of a Population of the Red-backed Shrike]." Journal fur Ornithologie 124:29-46.
Kempenaers, B. (1994) "Polygyny in the Blue Tit: Unbalanced Sex Ratio and Female Aggression Restrict Mate Choice." Animal Behavior 47:943-57.
--- (1993) "A Case of Polyandry in the Blue Tit: Female Extra-Pair Behavior Results in Extra Male Help." Ornis Scandinavica 24:246-49.
Kempenaers, B., G. R. Verheyen, and A. A. Dhondt (1997) "Extrapair Paternity in the Blue Tit (Parus caeruleus): Female Choice, Male Characteristics, and Offspring Quality." Behavioral Ecology 8:481-92.
--- (1995) "Mate Guarding and Copulation Behavior in Monogamous and Polygynous Blue Tits: Do Males Follow a Best-of-a-Bad-Job Strategy?" Behavioral Ecology and Sociobiology 36:33-42.
Krieg, D. C. (1971) The Behavioral Patterns of the Eastern Bluebird (Sialia sialis). New York State Museum and Science Service Bulletin no. 415. Albany: University of the State of New York.
Massa, R., L. Bottoni, L. Fornasari, and N. Sacchi (1995) "Studies on the Socio-Sexual and Territorial System of the Red-backed Shrike." Proceedings of the Western Foundation of Vertebrate Zoology 6:172-75.
Meek, S. B., R. J. Robertson, and P. T. Boag (1994) "Extrapair Paternity and Intraspecific Brood Parasitism in Eastern Bluebirds Revealed by DNA Fingerprinting." Auk 111:739-44.
* Owen, J. H. (1946) "The Eggs of the Red-backed Shrike." Oologists' Record 20:38-43.
* Pinkowski, B.C. (1977) "'Courtship Feeding' Attempt Between Two Male Eastern Bluebirds." Jack-Pine Warbler 55:45-46.
* Pounds, H. E. (1972) "Two Red-backed Shrikes Laying in One Nest." British Birds 65:357-58.
* Saetre, G.-P., and T. Slagsvold (1993) "Evidence for Sex Recognition from Plumage Color by the Pied Flycatcher, Ficedula hypoleuca." Animal Behavior 44:293-99.
* Slagsvold, T., and G.-P. Saetre (1991) "Evolution of Plumage Color in Male Pied Flycatchers (Ficedula hypoleuca): Evidence for Female Mimicry." Evolution 45:910-17.
Stanback, M. T., and W. D. Koenig (1992) "Cannibalism in Birds." In M.A. Elgar and B. J. Crespi, eds., Cannibalism: Ecology and Evolution Among Diverse Taxa, pp. 277-98. Oxford: Oxford University Press.
* Zeleny, L. (1976) The Bluebird: How You Can Help Its Fight for Survival. Bloomington: Indiana University Press.
{598}
SPARROWS, BLACKBIRDS, AND CROWS
 Weaverbirds
Weaverbirds
GRAY-CAPPED SOCIAL WEAVER (Pseudonigrita arnaudi)
IDENTIFICATION: A grayish buff, sparrow-sized bird with a pale gray-white crown. DISTRIBUTION: Northeast Africa. HABITAT: Bush, acacia savanna. STUDY AREA: Olorgesailie National Prehistoric Site, Kenya; subspecies P.a. arnaudi.
SOCIABLE WEAVER (Philetairus socius)
IDENTIFICATION: A drab, sparrowlike bird with brownish gray plumage and a black throat patch. DISTRIBUTION: Southwestern Africa. HABITAT: Scrub, savanna. STUDY AREA: University of California — Los Angeles.
RED BISHOP BIRD (Euplectes orix)
ORANGE BISHOP BIRD (Euplectes franciscanus)
IDENTIFICATION: Small, sparrowlike birds with black and brown plumage and various scarlet, red, or reddish orange patches on the chest, nape, crown, and rump. DISTRIBUTION: Sub-Saharan Africa. HABITAT: Moist grassland. STUDY AREAS: Several locations in South Africa, including near Cape Town, Howick, and Bloemfontein; University of Cape Town and University of Bielefeld.
{599}
Social Organization
Weaverbirds are named for the intricate — and often colossal — nests they weave. Sociable Weavers build giant condominium-like structures containing many nest chambers; each chamber may house up to 5 birds, while the entire colony can contain as many as 500 birds. Colonies of Gray-capped Social Weavers consist of family groups that each build several hanging nests (one is used as an actual nesting chamber, the others for sleeping). Several groups occupy the same tree, each situated in its own cluster. In both Gray-caps and Sociable Weavers, breeding birds form pair-bonds, and "helper" birds may assist them in their parenting duties. Adult male Bishop Birds establish territories during the breeding season on which they weave elaborate nests and court potential mates. The breeding system of Bishop Birds is polygynous: males mate with numerous females, each of whom lays eggs in one of the nests on his territory, but the male himself does not participate in any parental duties. Outside of the breeding season, Bishop Birds typically socialize in large, often mixed-species flocks.

A male Red Bishop Bird displaying fluffed "bumblebee" plumage, used during courtships with same- and opposite-sex partners
Description
Behavioral Expression: Male Gray-capped Social Weavers and Sociable Weavers sometimes participate in homosexual copulations. In Gray-caps, birds who participate in same-sex mounting are usually heterosexually paired males, often high-ranking individuals living in the same colony (tree). One male may fly from his home group in the tree to another male's residence, inviting him to mate by holding his body horizontal, raising his head and tail upward, and drooping and vibrating his wings. The other male then mounts him and proceeds to engage in a complete copulation sequence, probably including cloacal (genital) contact. Mounting between males occurs in Sociable Weavers as well, and about 9 percent of such mounts are also full copulations in which the mounted male crouches, quivers his wings, and moves his tail to the side as the mounting male lowers his tail (probably to achieve genital contact). Usually higher-ranking males mount lower-ranking ones, but all males both mount and are mounted by other males (to varying degrees). In addition, some males appear to form "companionships" with each other. Although males do not generally permit other males to roost with them in their nesting chambers, occasionally two males regularly associate with one another and sleep together in the same nest. This can occur even if one of the males is already paired to a female, in which case all three birds occupy the nest together. The two males may remain together for years, even cooperating in attacking other birds together. Some male "companions" participate in mountings and copulations with each other, but in other cases males do not seem to preferentially engage in sexual behavior with their companion.
On their nesting territories, male Red Bishop Birds court both females and males, the latter usually being younger males who have brown plumage (as do females). Courtship consists of two components: a flying display and a perched display. When a young male appears near his territory, an adult male approaches in a distinctive BUMBLE-FLIGHT, in which he fluffs up all of his plumage and flies with slow wingbeats, resembling a bumble-bee. The bird's bright red back or epaulet {600}
feathers are prominently displayed, and the bumble-flying may be accompanied by singing or a distinctive noise made by his flapping wings. Sometimes males bumble-fly toward another adult male rather than a younger male. After the display flight, the adult male lands near the other male and begins the SWIVEL DISPLAYS: he hops and twists around his perch, approaching the object of his attentions with ruffled plumage and a rattling call — a continuous stream of notes that sounds like zik-zik-zik. No homosexual copulations have been observed in this species in the wild, but heterosexual matings are not commonly seen either. In captivity Orange Bishop Birds — both adults and younger males — sometimes do try to mount younger males (including males of the closely related Golden Bishop Bird, Euplectes afer). In a homosexual encounter, one male approaches the other while pumping his body up and down and ruffling his feathers, followed by an attempt to copulate with the other male (who usually rejects his advances).
Frequency: In Sociable Weavers studied in captivity, as much as three-quarters of all mounting activity is between males, and three out of five full copulations are homosexual. The occurrence of same-sex mounting in wild birds of this species is not known, but its prevalence is probably comparable (especially since heterosexual activity is also apparently infrequent). In one study of wild Gray-capped Social Weavers, all sexual activity that was observed took place between males. In Red Bishops, approximately 6 percent of courtship bumble-flights are directed by one adult male toward another adult male; courtship of younger males probably occurs more often than this.
Orientation: Male Gray-capped Social Weavers that copulate with other males also mate and pair with females (and in fact may engage in homosexual activity while they are heterosexually paired). The same is true for some Sociable Weavers, although most males in this species mount both males and females, even if they are not paired. However, relatively few males participate in full heterosexual copulations, and those that do appear to have higher rates of homosexual activity as well. To the extent that they court or attempt to mount both males and females, some male Bishops are also bisexual. However, the males they pursue are usually indifferent, at best, to their approaches, indicating perhaps a more heterosexual orientation on their part.
Nonreproductive and Alternative Heterosexualities
In Gray-capped Social Weavers (and occasionally in Sociable Weavers), nonbreeding birds often help heterosexual pairs build their nest and feed their young. Some of these "helpers" are the pairs' young from a previous season who are delaying their own reproductive careers, while others are fully adult birds (who help out in about 18 percent of all feedings). {601}
Some nonbreeding youngsters, however, do not help their parents. Sociable Weavers may participate in nonmonogamous heterosexual mountings (in addition to the homosexual matings described above). Although most birds are probably faithful to their partners, some males in captivity have been observed mounting and copulating with females other than their mates. Female Red Bishops occasionally refuse to allow a male to mount them, vigorously pecking and displaying threat postures to repel him. In addition, males often court Bishops of other species, who are not usually attracted by such interspecies displays. Finally, female Red Bishops have been observed cannibalizing both their own and others' nests, eating some or all of the eggs.
Other Species
Adult male Red-shouldered Widowbirds (Euplectes axillaris) also sometimes court younger males.
Sources
(* asterisked references discuss homosexuality/transgender)
* Collias, E. C., and N. E. Collias (1980) "Individual and Sex Differences in Behavior of the Sociable Weaver Philetairus socius." In D. N. Johnson, ed., Proceedings of the Fourth Pan-African Ornithological Congress (Seychelles, 1976), pp. 243-51. Johannesburg: Southern African Ornithological Society.
* --- (1978) "Nest Building and Nesting Behavior of the Sociable Weaver Philetairus socius." Ibis 120:1-15.
* Collias, N. E., and E. C. Collias (1980) "Behavior of the Gray-capped Social Weaver (Psuedonigrita arnaudi) in Kenya." Auk 97:213-26.
Craig, A. J. F. K. (1982) "Mate Attraction and Breeding Success in the Red Bishop." Ostrich 53:246-48.
* --- (1980) "Behavior and Evolution in the Genus Euplectes." Journal fur Ornithologie 121:144-61.
* --- (1974) "Reproductive Behavior of the Male Red Bishop Bird." Ostrich 45:149-60.
Craig, A. J. F. K., and A. J. Manson (1981) "Sexing Euplectes Species by Wing-Length." Ostrich 52:9-16.
Maclean, G. L. (1973) "The Sociable Weaver." Ostrich 44:176-261.
Roberts, C. (1988) "Little Bishop Birds (Euplectes orix) in a Lafia Garden — Tom, Dick, Harry, and Fred." Nigerian Field 53:11-22.
Skead, C. J. (1959) "A Study of the Redshouldered Widowbird Coliuspasser axillaris axillaris (Smith)." Ostrich 30:13-21.
--- (1956) "A Study of the Red Bishop." Ostrich 27:112-26.
Woodall, P. F. (1971) "Notes on a Rhodesian Colony of the Red Bishop." Ostrich 42:205-10.
{602}
 Sparrows, Blackbirds, and Starlings
Sparrows, Blackbirds, and Starlings
HOUSE SPARROW (Passer domesticus)
BROWN-HEADED COWBIRD (Molothrus ater)
IDENTIFICATION: The familiar black-bibbed sparrow; Cowbird is iridescent black with a dark brown head. DISTRIBUTION: Throughout most of North and South America, Eurasia (House Sparrow); North and Central America (Cowbird). HABITAT: Woodland, prairie, farmland, human habitation. STUDY AREAS: Near Stillwater, Oklahoma, and Long Island, New York; subspecies M.a. ater and M.a. artemisiae.
WATTLED STARLING (Creatophora cinerea)
IDENTIFICATION: Light gray plumage, black wings and tail, and (in some birds) bare yellow head and fleshy black wattles. DISTRIBUTION: Eastern and southern Africa. HABITAT: Savanna, grassland, woodland. STUDY AREAS: University of Mainz and in Nieder-Olm, Germany.
Social Organization
Wattled Starlings usually associate in small, nomadic flocks, although up to a thousand birds may gather together to pursue locust swarms. Similarly, breeding colonies may contain thousands of nests when locusts are available, but usually birds nest in smaller groups containing a maximum of 400 pairs. Most individuals form monogamous pair-bonds, as do House Sparrows (who also generally nest in colonies). Brown-headed Cowbirds have a highly variable mating system: in many populations birds form (usually monogamous) pair-bonds, while in others they are promiscuous or form polygamous bonds with several individuals.

A male Brown-headed Cowbird in the "head-down" posture (right) inviting a male House Sparrow to mount him
Description
Behavioral Expression: Male Brown-headed Cowbirds sometimes solicit homosexual copulations from male House Sparrows. Cowbirds commonly invite birds of other species to preen them, but occasionally an interspecies encounter includes homosexual mounting when a House Sparrow is involved. This extraordinary behavior typically begins with a male Cowbird adopting a characteristic {603}
HEAD-DOWN posture next to a Sparrow, in which he bows his head, touching his lower bill to his breast feathers while crouching slightly and raising his wings a bit at the shoulders. The House Sparrow then mounts the Cowbird, grasping his head feathers in his beak while attempting to copulate. If he shows signs of leaving or lack of interest after a single mounting, the Cowbird will immediately resume the invitation posture next to him, insistently nudging the Sparrow with his head and persistently following him until he mounts again. This may continue for an extended time, with repeated homosexual mountings (five or more) occurring in a single session.
Homosexual courtships occasionally occur in Wattled Starlings. Males sometimes select another male as the object of their attentions, displaying to him with a number of stylized postures. Among these are the LATERAL DISPLAY, in which the male turns sideways and lets his wings hang down at his side (exposing their white feathers); the FRONTAL DISPLAY, where the courting male fluffs up his belly and back feathers, raising and quivering his wings while spreading his tail; and the distinctive VULTURE POSTURE, in which he stretches his entire body vertically while puffing out his breast feathers and tightly folding his wings against his sides like a vulture. A form of plumage transvestism also occurs in this species, in which some females develop a male appearance. Most males have a special seasonal nuptial plumage, growing two pendulous wattles from either side of their beaks and losing most of the feathers from their head, thereby exposing the yellow or black skin and two fleshy, comblike growths on the forehead. This feather loss has even been described as a form of "male pattern baldness" akin to the type of hair loss found in humans, and indeed it is regulated by male hormones (as is human baldness). While the majority of females never exhibit these plumage characteristics, a few females do acquire a male appearance with feather loss, wattles, and combs.
Frequency: Brown-headed Cowbirds regularly perform the head-down display toward other species in the wild, and approximately 36 percent of such displays are directed by male Cowbirds toward male House Sparrows; however, Sparrows respond with homosexual mounting probably only sporadically. Similarly, homosexual courtship is in all likelihood only an occasional occurrence in Wattled Starlings. About 2-10 percent of female Wattled Starlings are transgendered, exhibiting fairly complete wattles and/or baldness. Other females appear to fall along a continuum of plumage characteristics, with some individuals showing only partial wattle development or incomplete baldness.
{604}
Orientation: In all three of these species, not enough is yet known about the life histories of individuals participating in homosexual activity to determine their overall sexual orientation profiles. However, at least some male Wattled Starlings appear to preferentially select other males to court.
Nonreproductive and Alternative Heterosexualities
Although most heterosexual Wattled Starlings form monogamous pairs, occasionally males court and mate with females other than their mate. This may result in a female raising her young as a single parent if she is not paired to the male she mates with. The male sexual cycle is especially pronounced in this species, signaled by the seasonal development of wattles and baldness. House Sparrows often engage in multiple copulations during the same mating bout: a male may mount a female and achieve genital contact up to 30 times in rapid succession. In addition, promiscuous matings are quite common in this species: more than a quarter of all nests contain at least one chick sired by a bird other than its mother's mate. Some of these are the result of forced matings that occur during COMMUNAL DISPLAYS, in which "gangs" of up to ten males chase a female, peck at her genitals, and try to mount her. Such displays and the associated sexual activity often take place during nonfertilizable periods as well. Courting male Cowbirds also frequently harass females, and on average only about 12 percent of heterosexual consortships in this species culminate in copulation. In pair-bonding populations of Brown-headed Cowbirds, about 16 percent of courtships are actually between birds not paired to one another, and some promiscuous matings occur as well. In both this species and House Sparrows, a few pairs switch mates during the breeding season, and a subset of birds (about 5-6 percent) are polygamous.
Large numbers of male Cowbirds are nonbreeders: more than half of all males in some populations are unpaired, and only a third of males actually copulate with females in some years. Brown-headed Cowbirds are also BROOD PARASITES, which means that females always lay their eggs in the nests of other bird species and take no part in raising their own young. Infanticide occurs in 9-12 percent of House Sparrow nests, often when a female who has lost her mate pairs with a new male (who pecks her young to death in order to father his own offspring). Females in polygamous trios also occasionally kill one another's nestlings. Sometimes, however, a female whose mate has been replaced by an infanticidal male will stop laying eggs (by interrupting or delaying ovulation) in order not to lose any more young, and some replacement males adopt rather than kill their mate's young.
Other Species
Male Sharp-tailed Sparrows (Ammodramus caudacutus), a North American species, sometimes mount other males. Adolescent male Yellow-rumped Caciques (Cacicus cela), a South American blackbird, frequently mount fledglings of both sexes. Sexual behavior toward these younger birds is usually part of an overall pattern of harassment, in which the adolescent males (often in groups) chase, peck, attack, and sometimes even knock fledglings from their perches (often resulting in {605}
their death by drowning if they fall into water). About 36 percent of such harassments (and the associated sexual behavior) involve same-sex interactions.
Sources
(* asterisked references discuss homosexuality/transgender)
Craig, A. J. F. K. (1996) "The Annual Cycle of Wing Moult and Breeding in the Wattled Starling Creatophora cinerea." Ibis 138:448-54.
Darley, J. A. (1978) "Pairing in Captive Brown-headed Cowbirds (Molothrus ater)." Canadian Journal of Zoology 56:2249-52.
* Dean, W. R. J. (1978) "Plumage, Reproductive Condition, and Moult in Non-Breeding Wattled Starlings." Ostrich 49:97-101.
Friedmann, H. (1929) The Cowbirds: A Study in the Biology of Social Parasitism. Springfield, Ill.: Charles C. Thomas.
* Greenlaw, J. S., and J. D. Rising (1994) "Sharp-tailed Sparrow (Ammodramus caudacutus)." In A. Poole and F. Gill, eds., The Birds of North America: Life Histories for the 21 st Century, no. 112. Philadelphia: Academy of Natural Sciences; Washington, D.C.: American Ornithologists' Union.
* Griffin, D. N. (1959) "Apparent Homosexual Behavior Between Brown-headed Cowbird and House Sparrow." Auk 76:238-39.
* Hamilton, J. B. (1959) "A Male Pattern Baldness in Wattled Starlings Resembling the Condition in Man." Annals of the New York Academy of Sciences 83:429-47.
Laskey, A. R. (1950) "Cowbird Behavior." Wilson Bulletin 62:157-74.
Liversidge, R. (1961) "The Wattled Starling (Creatophora cinerea [Menschen])." Annals of the Cape Provincial Museums 1:71-80.
Lowther, P. E. (1993) "Brown-headed Cowbird (Molothrus ater)." In A. Poole, P. Stettenheim, and F. Gill, eds., The Birds of North America: Life Histories for the 21st Century, no. 47. Philadelphia: Academy of Natural Sciences; Washington, D.C.: American Ornithologists' Union.
Lowther, P. E., and C. L. Cink (1992) "House Sparrow (Passer domesticus)." In A. Poole, P. Stettenheim, and F. Gill, eds., The Birds of North America: Life Histories for the 21 st Century, no. 12. Philadelphia: Academy of Natural Sciences; Washington, D.C.: American Ornithologists' Union.
Moller, A. P. (1987) "House Sparrow, Passer domesticus, Communal Displays." Animal Behavior 35:203-10.
* Robinson, S. K. (1988) "Anti-Social and Social Behavior of Adolescent Yellow-rumped Caciques (Icteri-nae: Cacicus cela)." Animal Behavior 36:1482-95.
Rothstein, S. I. (1980) "The Preening Invitation or Head-Down Display of Parasitic Cowbirds: II. Experimental Analysis and Evidence for Behavioral Mimicry." Behavior 75:148-84.
Rothstein, S. I., D.A. Yokel, and R. C. Fleischer (1986) "Social Dominance, Mating and Spacing Systems, Female Fecundity, and Vocal Dialects in Captive and Free-Ranging Brown-headed Cowbirds." Current Ornithology 3:127-85.
Scott, T. W., and J. M. Grumstrup-Scott (1983) "Why Do Brown-headed Cowbirds Perform the Head-Down Display?" Auk 100:139-48.
* Selander, R. K., and C. J. La Rue, Jr. (1961) "Interspecific Preening Invitation Display of Parasitic Cowbirds." Auk 78:473-504.
* Sontag, W.A., Jr. (1991) "Habitusunterschiede, Balzverhalten, Paarbildung, und Paarbindung beim Lap-penstar Creatophora cinerea [Behavior Differences, Courtship, Pair Formation, and Pair Bonding in the Wattled Starling]." Acta Biologica Benrodis 3:99-114.
--- (1978/79) "Remarks Concerning the Social Behavior of Wattled Starlings, Creatophora cinerea (Menschen)." Journal of the Nepal Research Center 2/3:263-68.
Teather, K. L., and R. J. Robertson (1986) "Pair Bonds and Factors Influencing the Diversity of Mating Systems in Brown-headed Cowbirds." Condor 88:63-69.
Uys, C. J. (1977) "Notes on Wattled Starlings in the Western Cape." Bokmakierie 28:87-89.
Veiga, J. P. (1993) "Prospective Infanticide and Ovulation Retardation in Free-Living House Sparrows." Animal Behavior 45:43-46.
--- (1990) "Infanticide by Male and Female House Sparrows." Animal Behavior 39:496-502.
Wetton, J. H., and D.T. Parkin (1991) "An Association Between Fertility and Cuckoldry in the House Sparrow, Passer domesticus." Proceedings of the Royal Society of London, Series B 245:227-33.
Yokel, D. A. (1986) "Monogamy and Brood Parasitism: An Unlikely Pair." Animal Behavior 34:1348-58.
Yokel, D. A., and S. I. Rothstein (1991) "The Basis for Female Choice in an Avian Brood Parasite." Behavioral Ecology and Sociobiology 29:39-45.
{606}
 Crows
Crows
BLACK-BILLED MAGPIE (Pica pica)
IDENTIFICATION: A pigeon-sized, crowlike bird with striking iridescent, black-and-white plumage and a long, greenish purple tail. DISTRIBUTION: Eurasia, North Africa, western North America. HABITAT: Woodland, scrub, grassland, savanna. STUDY AREAS: Haren, the Netherlands; University of Groningen, the Netherlands; subspecies P.p. pica.
(Eurasian) JACKDAW (Corvus monedula)
IDENTIFICATION: A small crow with black plumage and gray on the back of the head. DISTRIBUTION: Eurasia, North Africa. HABITAT: Forest, grassland, farmland. STUDY AREAS: Haren, the Netherlands; Max-Planck Institute; subspecies C.m. spermologus.
(Common) RAVEN (Corvus corax)
IDENTIFICATION: A large (2 foot), all-black bird similar to a crow but much bulkier. DISTRIBUTION: Eurasia, North America. HABITAT: Varied, including forest, plains, desert. STUDY AREA: Max-Planck Institute; subspecies
C.c. corax.
Social Organization
All three of these Crow species are quite gregarious, often associating in flocks and communal roosts. Individuals generally form long-term mated pairs, and Jackdaws usually nest in colonies. Magpies sometimes participate in remarkable group displays known as CEREMONIAL GATHERINGS, noisy aggregations that may be related to territory acquisition.
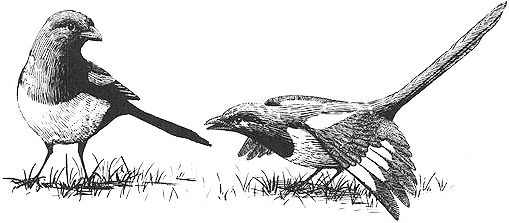
Homosexual courtship in Black-billed Magpies: "tilting" (left) and "begging"
Description
Behavioral Expression: Black-billed Magpies sometimes court and form pair-bonds with birds of the same sex. Partners include adult males with younger or juvenile males (less than one year old), or else two females or two males of the same age, usually juveniles but also sometimes two adult males. A typical homosexual courtship — for example, between two males — begins with one bird ritually BEGGING the other by crouching in front of him and flapping or quivering his wings {607}
while uttering a begging call. The other male responds by hopping in tight circles around him, fluffing up his white feathers and flicking his wings; he may also BABBLE-SING, a varied combination of warbling, chattering, and yelping notes. The circling male often adopts a TILTING posture, in which he points his head and tail sideways toward the other male. If the male he is courting flies off, a courtship pursuit known as CHASE-HOPPING may develop, in which one male follows the other while alternately flying and hopping. Sometimes the two birds also HOVER-FLY, one in front of the other, using a rhythmic, undulating flight pattern. The same series of behaviors is seen in courtships between females (as well as in heterosexual interactions). Homosexual courtship sessions can last for up to half an hour. After courting, two birds of the same sex may form a pair-bond. Paired birds stay near each other, follow one another, and often cooperate in evicting intruders from their territory. They also frequently sit close together and preen one another or engage in mutual BILLING, in which they affectionately nibble at each other's beak. Sometimes, homosexual mates also pull or nibble on the same leaf or twig and pass it back and forth between them; this is known as TUGGING. Same-sex pair-bonds are generally of shorter duration than adult heterosexual bonds and last from a few days to several months. However, adult males sometimes form longer-lasting homosexual pairs, and the two birds may even build a nest together (which typically takes five to seven weeks to construct).
Female Jackdaws occasionally develop pair-bonds with other females. In some cases, an older female pairs with a younger one, and the two build a nest together even though the juvenile bird is too young to lay eggs. Later, they might construct a unique "double nest" consisting of two adjacent cups and lay infertile eggs in both cups. Sometimes, a homosexual pair is joined by a male, who may bond with one or both of the females to form a bisexual trio; in this way, the females can lay fertilized eggs. However, they are often unable to successfully care for their offspring, precisely because their bond to each other means that they try to stay together all the time. The two females incubate their eggs and brood their youngsters simultaneously, each sitting on one cup. When the male arrives for his shift, however, they both depart together, leaving the male to try to cover and protect both sets of eggs or nestlings at the same time (which he is usually unable to do). Sometimes, a {608}
bi-sexual trio forms when a female joins a heterosexual pair and develops a strong bond with the female partner. The two females engage in courtship and pair-bonding activities such as mutual preening or COURTSHIP-FEEDING, in which one partner begs the other by crouching, fluttering her wings, and quivering her tail. Both females might mate with the male and lay fertile eggs, although the bond between them can end up being stronger than the original heterosexual bond. In fact, in one case the female partners were not able to properly care for their young because the male denied the female "interloper" access to the nestlings. Homosexual bonds also sometimes develop between widowed and nonbreeding females. In these cases, females that lose their male partners during the breeding season may attract unmated females to pair with them; some of these widows are mothers, while others have no offspring. Unlike bisexual trios, which may remain together for years, these female bonds appear to be more transitory, usually lasting only for several weeks until the breeding season is over.
Female homosexual pairs also occasionally occur in Ravens, including incestuous bonds between yearling sisters. Birds in same-sex pairs engage in intense courtship activity similar to heterosexual pairs, such as mutual preening and courtship-feeding.
Frequency: Among Jackdaws in the wild, approximately 5 percent of trios include bonding between the two female partners, while about 10 percent of widowed females form homosexual pairs. Overall, though, same-sex bonds probably represent no more than 1 percent of all pairs/trios. Homosexual activity occurs sporadically in Black-billed Magpies and Ravens as well: about 1 percent of nesting pairs of Magpies, for example, consist of two males. Although homosexual copulations between Magpies have not yet been recorded, heterosexual matings are also infrequently observed (only 9 male-female copulations, for example, were recorded during one 300 — hour study period).
Orientation: Homosexual behavior is most prevalent among younger Black-billed Magpies, some of whom also court members of the opposite sex; many of these birds go on to form heterosexual attachments when they become adults, but most are nonbreeders during the time of their same-sex activities. Some adult males, however, continue to court younger males or pair with other adult males. In Jackdaws and Ravens many different forms of bisexuality occur. Some females are simultaneously bonded to both males and females, and such trios may develop out of either an initial heterosexual or homosexual pair-bond. Some females are sequentially bisexual, developing same-sex bonds only after losing their male partners. However, those Jackdaw females who enter into homosexual pairs as nonbreeders may have no prior (and perhaps no subsequent) heterosexual experience.
Nonreproductive and Alternative Heterosexualities
Black-billed Magpies and Jackdaws participate in a number of nonprocreative heterosexual activities. Birds sometimes copulate long after fertilization has taken {609}
place — even during incubation or chick-rearing — while courtship and mounting may occur well in advance of the breeding season in Magpies. In both species, birds form pair-bonds as juveniles, long before they are able to breed. Several alternatives to the monogamous pair and nuclear family are also to be found in these species of Crows. In Black-billed Magpies, for example, courtship and copulations with birds outside of the pair-bond can be more frequent than within-pair matings. In addition, some pairs get divorced: about half of all males and just under two-thirds of females change mates, and some may pair with up to three different partners over the course of their lives. Other Magpies, however, do remain in lifelong, faithful pair-bonds. About 6-10 percent of adult Jackdaw pairs, and a third of all juvenile pairs, get divorced. Polygamous trios also sometimes develop in Magpies (1-2 percent of bonds), but are fairly common in Jackdaws, comprising about 14 percent of all bonds. These usually result from an unpaired female joining an existing male-female pair; unlike the bisexual trios described above, she usually bonds only with the male of the pair, not the female. Occasionally, she may oust the former female and start a new family with the male; frequently, however, the newcomer female does not breed even if she remains in the trio for several years.
Nonbreeding couples also occur in Jackdaws and Ravens, as well as a significant population of single, nonreproducing birds. The latter are found in Magpies as well, where they make up 20-60 percent of the population and may be solitary or form their own flocks. In addition, about half of all Magpies that survive to adulthood leave no descendants (even if they try to breed). Some nonbreeding Jackdaw pairs — or couples who have tried unsuccessfully to breed — end up harassing breeding pairs by invading their nests, fighting with them, and even attacking and occasionally killing their young with vicious pecks. Nearly a third of all breeding pairs in some populations are harassed in this way. Cannibalism of young and eggs by neighboring breeding pairs also sometimes occurs in Jackdaws. Magpies and Ravens occasionally adopt unrelated youngsters when they pair with a bird that has lost its mate. Magpie families also sometimes foster-parent chicks as a result of the extraordinary phenomenon of "egg transfer": in some populations, up to 8 percent of nests contain a foreign Magpie egg, but not as a result of being laid there. Rather, another Magpie has physically carried its own egg in its beak to the new nest, placing it there to hatch and be raised by the host parents. Stealing and cannibalism of young or eggs has also been reported among Black-billed Magpies: about 7 percent of nests are robbed by other Magpies. In addition, at least 30 percent of nestlings die of starvation as a result of competition from their siblings, or from direct attacks or cannibalism by their nest-mates.
Other Species
Homosexual courtship occurs in some species of Jays (closely related to Crows), so far observed only in captivity. Male Gray-breasted or Mexican Jays (Aphelocoma ultramarina), a species found in the American Southwest and Mexico, have been seen COURTSHIP-FEEDING younger males in mixed-sex groups. This behavior — also part of heterosexual courtships — involves one male offering the other a food item; the latter accepts it while quivering his wings, crouching, and uttering {610}
a low kwa kwa kwa call. After eating the food or storing it in a cache, he may follow the other male to repeat the episode. Female San Bias Jays (Cyanocorax san-blasianus) of Mexico have been known to court other females in same-sex groups in captivity, using the SOTTO VOCE SONG DISPLAY. While making soft, throaty vocalizations, one female sidles up to the other on a perch with her tail raised and belly feathers fluffed.
Sources
(* asterisked references discuss homosexuality/transgender)
Antikainen, E. (1981) "The Breeding Success of the Jackdaw Corvus monedula in Nesting Cells." Ornis Fennica 58:72-77.
* Baeyens, G. (1981a) "Magpie Breeding Success and Carrion Crow Interference." Ardea 69:125-39.
--- (1981b) "Functional Aspects of Serial Monogamy: The Magpie Pair-Bond in Relation to Its Territorial System." Ardea 69:145-66.
* --- (1979) "Description of the Social Behavior of the Magpie (Pica pica)." Ardea 67:28-41.
Birkhead, T. (1991) The Magpies: The Ecology and Behavior of Black-billed and Yellow-billed Magpies. London: T. and A. D. Poyser.
Birkhead, T., and J. D. Biggins (1987) "Reproductive Synchrony and Extra-Pair Copulation in Birds." Ethology 74:320-34.
Birkhead, T., S. F. Eden, K. Clarkson, S. F. Goodburn, and J. Pellatt (1986) "Social Organization of Magpies Pica pica." Ardea 74:59-68.
Buitron, D. (1988) "Female and Male Specialization in Parental Care and Its Consequences in Black-billed Magpies." Condor 90:29-39.
--- (1983) "Extra-Pair Courtship in Black-billed Magpies." Animal Behavior 31:211-20.
Coombs, F. (1978) The Crows: A Study of the Corvids of Europe. London: B. T. Batsford.
Dhindsa, M. S., and D. A. Boag (1992) "Patterns of Nest Site, Territory, and Mate Switching in Black-billed Magpies (Pica pica).' Canadian Journal of Zoology 70:633-40.
Dunn, P. O., and S. J. Hannon (1989) "Evidence for Obligate Male Parental Care in Black-billed Magpies." Auk 106:635-44.
* Hardy, J. W. (1974) "Behavior and Its Evolution in Neotropical Jays (Cissilopha)." Bird-Banding 45:253-68.
* --- (1961) "Studies in Behavior and Phylogeny of Certain New World Jays (Garrulinae)." University of Kansas Science Bulletin 42:13-149.
Jerzak, L. (1995) "Breeding Ecology of an Urban Magpie Pica pica Population in Zielona Gora (SW Poland)." Acta Ornithologica 29:123-33.
* Lorenz, K. (1972) "Pair-Formation in Ravens." In H. Friedrich, ed., Man and Animal: Studies in Behavior, pp. 17-36. New York: St. Martin's.
* --- (1935) "Der Kumpan in der Umwelt des Vogels." Journal fur Ornithologie 83:10-213, 289-413. Reprinted as "Companions as Factors in the Bird's Environment." In K. Lorenz (1970) Studies in Animal and Human Behavior, vol.1, pp. 101-258. Cambridge, Mass.: Harvard University Press.
Ratcliffe, D. (1997) The Raven: A Natural History in Britain and Ireland. London: T. and A. D. Poyser.
Reynolds, P. S. (1996) "Brood Reduction and Siblicide in Black-billed Magpies (Pica pica)." Auk 113:189-99.
* Roell, A. (1979) "Bigamy in Jackdaws." Ardea 67:123-29.
--- 1978) "Social Behavior of the Jackdaw, Corvus monedula, in Relation to Its Niche." Behavior 64:1-124.
Trost, C. H., and C. L. Webb (1986) "Egg Moving by Two Species of Corvid." Animal Behavior 34:294-95.
{611}
BIRDS OF PARADISE, BOWERBIRDS, AND OTHERS
 Birds of Paradise
Birds of Paradise
RAGGIANA'S BIRD OF PARADISE (Paradisaea raggiana)
IDENTIFICATION: A crow-sized bird; male has a bright yellow head, an iridescent green throat, and a long "tail" of orange flank feathers; female has a duller yellow head and brown facial mask and no orange "tail." DISTRIBUTION: Southern and northeastern Papua New Guinea. HABITAT: Lowland and hill forests. STUDY AREA: In captivity at the Baiyer River Sanctuary, Mt. Hagen, Papua New Guinea; subspecies P.r. augustaevictoriae.
VICTORIA'S RIFLEBIRD (Ptiloris victoriae)
IDENTIFICATION: Adult males are black with an iridescent sheen and metallic crown, throat, and central tail feathers; females and younger males are drabber, with brown, buff, and off-white plumage. DISTRIBUTION: Northeast Queensland, Australia. HABITAT: Rain forest, eucalyptus forest, swamp woodlands. STUDY AREAS: Southern Atherton Tableland, Queensland, Australia, including near Townsville and in the Ingham and Palmerston National Parks.
Social Organization
Male Raggiana's Birds of Paradise perform courtship displays on communal "courts" or LEKS (located in tree branches) in groups of up to eight individuals at a time; females visit the leks in pairs or small groups. Male Victoria's Riflebirds display singly. Both species have a polygamous or promiscuous mating system, in which males copulate with more than one female but do not participate in any parental duties.

A male Victoria's Riflebird courting another male with the "circular wings and gape" display
{612}
Description
Behavioral Expression: Male Victoria's Riflebirds sometimes court younger males with a series of spectacular displays (also seen in heterosexual courtships). Adult males attract individuals of both sexes to their display perches by repeatedly calling with a loud yass note and opening their bills to expose the bright yellow mouth interior. When an interested male (or female) approaches, the courting male begins his CIRCULAR WINGS AND GAPE display, in which he fans his wings to form a perfect circle with the tips meeting above his head while simultaneously raising and lowering his body. The wing circle is held vertically as he gapes his bill and presents his iridescent throat feathers to the other male. If the latter lands on the display perch, the courtship progresses to its final phase, the astounding ALTERNATE WINGS CLAP display. The adult male begins swaying and twisting his body from side to side, alternately raising and lowering each wing so that they "clap" with a dull, thudding sound when they meet above his head. Gradually the wings are brought forward while being clapped so that they encircle the younger male. The tempo of the display is increased until, at its climax, the wing-clapping reaches an astounding speed — each wing alternately encircles the observing male at a rate approaching twice a second. The courted male becomes mesmerized by the brilliant metallic bluish green throat shield of the displaying male as it is jerked violently back and forth, and he may respond by simultaneously performing his own alternate wings clap display. If both males are displaying, they alternate wing movements, gradually crouching while arching their heads back and quivering all over. They often perform several bouts of mutual display with brief rests between each. Occasionally one male briefly mounts the other; full copulation does not usually occur, though, since the mounted male often simply flies away from underneath the male mounting him.
Courtship displays between female Raggiana's Birds of Paradise involve some of the same postures and movements used in heterosexual courtship in this species. One female arches her wings above her back and {613}
then claps them against her sides, all the while bobbing up and down and erecting the feathers on her back and flanks as she dances in front of the other female. Every now and then she makes an ur, ur call, and she may even hang upside down on a branch while flicking her wings and calling. This dramatic pose is held for a minute or more at a time. Males use this inverted posture to show off their orange "tail" plumes during courtship, but females perform this display even though they do not have such ornate feathers. Sometimes two females engage in mutual display, facing each other while holding their wings stiffly above their backs and bobbing.
Frequency: Courtship displays between male Victoria's Riflebirds occur fairly often at some times of the year, especially during the postmolting period in February and March. Although homosexual mountings are not common in this species, opposite-sex matings have in fact been seen only a few times during more than a hundred courtship displays observed in the wild.
Orientation: At least some male Victoria's Riflebirds are functionally bisexual, courting and attempting to mate with both males and females. In Raggiana's Bird of Paradise, courtship display between females has so far only been observed in captivity in the absence of males, so this behavior may be the expression of a potential or "latent" bisexual capacity. More detailed field observations and life histories are required, though, before any definitive statements can be made regarding the sexual orientation of individuals that engage in this activity.
Nonreproductive and Alternative Heterosexualities
Only about 11 percent of lek visits by female Raggiana's Birds of Paradise actually result in heterosexual mating. More often than not, a courtship interaction is broken off by a female before copulation occurs. In addition, males often chase females and behave aggressively toward them on the lek, which deters them from participating in sexual interactions. When mating does take place, the female often mounts the male prior to copulation. Such REVERSE mounts are then followed by a display in which the male appears to pummel his partner with his wing. The female crouches on her perch absorbing the blows for 20-35 seconds, after which copulation occurs. Nonreproductive matings also take place outside of the breeding season. Many males may delay breeding up to five or more years once becoming sexually mature, since they do not generally acquire their ornate plumage (used in courtship displays) for several years.
Other Species
Male Greater Birds of Paradise (Paradisaea apoda), a species closely related to Raggiana's, also sometimes court and mount younger males.
Sources
(* asterisked references discuss homosexuality/transgender)
Beehler, B.M. (1989) "The Birds of Paradise." Scientific American 261(6):116-23.
--- (1988) "Lek Behavior of the Raggiana Bird of Paradise." National Geographic Research 4:343-58.
* Bourke, P. A., and A. F. Austin (1947) "The Atherton Tablelands and Its Avifauna." Emu 47:87-116. {614}
Davis, W. E., Jr., and B. M. Beehler (1994) "Nesting Behavior of a Raggiana Bird of Paradise." Wilson Bulletin 106:522-30.
* Frith, C. B. (1997) Personal communication.
--- (1981) "Displays of Count Raggi's Bird-of-Paradise Paradisaea raggiana and Congeneric Species." Emu 81:193-201.
* Frith, C. B., and W. T. Cooper (1996) "Courtship Display and Mating of Victoria's Riflebird Ptiloris victoriae with Notes on the Courtship Displays of Congeneric Species." Emu 96:102-13.
Frith, C. B., and D. W. Frith (1995) "Notes on the Nesting Biology and Diet of Victoria's Riflebird Ptiloris victoriae." Emu 95:162-74.
Gilliard, E. T. (1969) "Queen Victoria Rifle Bird" and "Count Raggi's Bird of Paradise." In Birds of Paradise and Bower Birds, pp. 112-17,222-29. Garden City, N.Y.: Natural History Press.
Lecroy, M. (1981) "The Genus Paradisaea — Display and Evolution." American Museum Novitates 2714:1-52.
* Mackay, M. (1981) "Display Behavior by Female Birds of Paradise in Captivity." Newsletter of the Papua New Guinea Bird Society 185/186 (November-December):5.
 Bowerbirds
Bowerbirds
REGENT BOWERBIRD (Sericulus chrysocephalus)
IDENTIFICATION: A thrush-sized bird (12 inches) with velvety black plumage and brilliant golden yellow crown, nape, upper back, and wing feathers. DISTRIBUTION: East-central Australia. HABITAT: Humid forests. STUDY AREA: Sarabah Range, Queensland, Australia.
Social Organization
Regent Bowerbirds have a polygamous or promiscuous mating system: males court and mate with multiple partners on display territories (see below). Following the mating season birds often associate in cosexual flocks of 10-20 individuals.

A bower constructed by a male Regent Bowerbird, used during courtship displays to both males and females
Description
Behavioral Expression: Male Regent Bowerbirds build elaborate structures called BOWERS, in which they court both males and females. Bowers built by adult {615}
males form an "avenue" on the ground consisting of two parallel walls of twigs — 10-12 inches high and 7-8 inches long — which are inserted into a platform of twigs on the ground. In some bowers the walls form a sort of arched walkway, while other bowers have triangular-shaped walls of different sizes. The bower is usually decorated with a dozen or so "display objects" or ornaments that are often selected for their color and strewn on the platform. These include brown snail shells, berries, green or purplish leaves, brown fruits and seeds, cicada husks, occasionally a yellow or pink flower petal, and even scavenged pieces of blue plastic. Remarkably, some birds also "paint" the walls of their bowers by applying, with their bills, macerated plant material mixed with saliva, giving some of the twigs a yellowish green coating. Adult males display in their bowers to females and younger males by flicking their wings and bowing their heads to show off their brilliant orange and yellow neck feathers. Sometimes they also ritually "present" display objects to the bird they are courting by picking up one of the ornaments from the avenue and holding it in their bill while facing their partner.
Juvenile males also build bowers and court both males and females. In some cases, they use the same bower shapes and displays as adults, but in other cases they utilize their own patterns, some of which may be unique to same-sex interactions. Some bowers built by younger males are more in the form of a horseshoe — open only at one end — with the sticks woven horizontally rather than vertically. When courtship-displaying to an adult male, the younger male enters the bower and squats with his tail toward the entrance. The adult male then rushes toward the juvenile, who is behind the closed end of the bower, or sometimes goes around to the entrance and tweaks the younger male's tail. When courting other juvenile birds of both sexes, young males may perform a dance in the center of the bower, picking up an ornament and bobbing up and down with half-opened wings, then tossing the object over the bower wall. The bird(s) watching this display sweep and brush the ground with their wings.
Many younger males are transvestite, having a plumage coloration that more closely resembles that of adult females than adult males. It takes males up to seven years to develop the full yellow, orange, and black feathers typical of adult males, {616}
and during that time many actually exhibit plumages that are intermediate between the adult male and female patterns.
Frequency: Among adult male Regent Bowerbirds, 15 percent of their bower courtship time is spent displaying to other males, while 28 percent of juvenile males' display time is same-sex.
Orientation: Regent Bowerbirds that court other males are probably bisexual, since they display to birds of both sexes.
Nonreproductive and Alternative Heterosexualities
A large segment of the male Regent Bowerbird population is nonbreeding: only about a third of all adult males maintain bowers, and of those that do, only a fraction actually mate with females. In addition, heterosexual courtship interactions rarely result in copulation: in only about 7 percent of female visits to bowers does mating actually take place, since the female often leaves while the male is displaying (and in 10 percent of female visits the male does not display at all).
Other Species
Adult male Satin Bowerbirds (Ptilonorhynchus violaceus) from Australia have also been observed performing courtship displays toward younger males.
Sources
(* asterisked references discuss homosexuality/transgender)
Chaffer, N. (1932) "The Regent Bird." Emu 32:8-11.
* Gilliard, E. T. (1969) "Australian Regent Bower Bird." In Birds of Paradise and Bower Birds, pp. 335-44. Garden City, N.Y.: Natural History Press.
Goddard, M. T. (1947) "Bower-Painting by the Regent Bower-bird." Emu 47:73-74.
* Lenz, N. (1994) "Mating Behavior and Sexual Competition in the Regent Bowerbird Sericulus chrysocephalus." Emu 94:263-72.
* Marshall, A. J. (1954) "Satin Bower-bird, Ptilonorhynchus violaceus (Vieillot)" and "Regent Bower-bird, Sericulus chrysocephalus (Lewin)." In Bower-birds: Their Displays and Breeding Cycles, pp. 26-71, 109-18. Oxford: Oxford University Press.
* Phillipps, R. (1905) "The Regent Bird (Sericulus melinus)." Avicultural Magazine (new series) 4:51-68, 88-96, 123-31.
Plomley, K. F. (1935) "Bower of the Regent Bower-bird." Emu 34:199.
{617}
 Lyrebirds
Lyrebirds
SUPERB LYREBIRD (Menura novaehollandiae)
IDENTIFICATION: A pheasant-sized bird with brownish gray plumage, powerful legs and claws, and long, ornate tail feathers. DISTRIBUTION: Southeastern Australia. HABITAT: Rain forest, eucalyptus, other forests. STUDY AREAS: Sherbrooke Forest near Melbourne, Australia; Adelaide Zoo, South Australia.
Social Organization
Adult male Superb Lyrebirds establish territories on which they build earth mounds three to five feet wide, used as platforms for courtship displays. The mating system is polygamous or promiscuous: birds mate with multiple partners without developing long-lasting pair-bonds, and males do not contribute to raising their offspring. Adolescent Lyrebirds often associate in small groups, sometimes consisting only of males.

A male Superb Lyrebird (foreground) courting a younger male in the forests of Australia. He is singing and performing the dramatic "full-face display," in which he fans his elaborate tail plumes forward over his head.
Description
Behavioral Expression: Male Superb Lyrebirds often court younger (adolescent) males on encountering them either in their groups or singly. The adult male closely follows the younger male, sometimes for hours at a time, periodically performing the WING-RAISING DISPLAY, in which he lifts and fans one wing toward his partner. With neck outstretched, he serenades the other male with a variety of extraordinary vocalizations, such as "whisper-song"; chortling; a call that can be rendered phonetically as clonk clonk clickety clickety click; another vocalization that sounds like the grinding of a pair of scissors; as well as uncanny imitations of other birds' songs. Sometimes the courtship progresses to spectacular displays of the male's elongated and stunningly beautiful tail feathers. In the dazzling FULL-FACE DISPLAY, for example, the older male arches his tail forward over his head and spreads the silvery filamentary feathers, hiding his body behind a gossamer fan framed by the elegant lyre-shaped, chestnut-striped outer tail feathers (for which the bird is named). Often the entire tail is vibrated to create a shimmering effect directed toward the other male. This may be followed by the INVITATION DISPLAY, in which the male almost "closes up" his quivering tail fan while still holding it forward (the feather tips almost touching the ground in front of him), all the while {618}
repeating a blick blick blick call. Occasionally, the younger male being courted by an adult is his own offspring. Adult males also sometimes mount adolescent males, even during the nonbreeding season when the adult bird is molting and has shed his elaborate tail plumes. Genital contact probably does not occur, though, since the younger male usually does not facilitate the interaction.
Adolescent male Lyrebirds also sometimes mount one another as well as particpate in homosexual courtship displays. Courtship between younger males is usually mutual, with both birds singing and displaying to one another. Occasionally an adult and an adolescent male also engage in mutual display, and adult females (in captivity) have been observed performing a similar courtship display to each other. Typically, the two adolescent males circle around each other on a display mound, tails raised in a fan shape with their feathers intermingled and beaks nearly touching while performing a vocal duet. One male may also perform a full-face or invitation display, and his partner sometimes runs underneath his outstretched tail the way a female does during heterosexual courtship. Sometimes the two males engage in what appears to be a form of COURTSHIP-FEEDING: in response to begging from his partner, one male regurgitates a worm or other food item as an offering to the other, who promptly eats it. This behavior appears to be unique to homosexual courtships. Two younger males often form a "companionship": in addition to courting each other, they follow one another, feed together (even {619}
digging in the same hole for food items), roost next to each other, and share bathing pools. These male pair-bonds usually last for only a few days, and adolescent males often form multiple serial attachments of this sort.

Two younger male Superb Lyrebirds performing a mutual courtship display
Frequency: Homosexual courtship occurs fairly often in Superb Lyrebirds. Adult males approach groups of adolescent males approximately once every three days during the breeding season and roughly once every day and a half outside the breeding season, and 93 percent of these encounters include courtship. In comparison, heterosexual encounters occur about four times as often as homosexual encounters during the breeding season, and about twice as often outside the breeding season. Adult males associate with adolescent males (in groups or singly) more than half of the time they are away from their display mounds. Same-sex mounting occurs less frequently than courtship interactions between males.

An adult male Superb Lyrebird (with molted tail feathers) mounting a younger male in Australia
Orientation: Most adult males are functionally bisexual, courting and mounting both females and younger males. Adolescent males appear to be more exclusively homosexual: most individuals form same-sex companionships and engage in homosexual courtship (and sometimes mounting) for several years prior to mating heterosexually. Some females probably have a bisexual potential that manifests itself in the absence of males (for example, in captivity).
Nonreproductive and Alternative Heterosexualities
Male and female Superb Lyrebirds lead largely separate lives. Other than brief encounters during the breeding season for courtship and copulation, the two sexes rarely interact: only about 8-10 percent of all Lyrebird sightings are of males and females together. Because males do not contribute at all to parenting, incubation and chick-raising often become burdensome for females — and by extension, potentially harmful for the eggs and/or chicks. During the early stages of incubation, females regularly leave their eggs unattended during the day for up to seven hours at a time to feed, causing the egg temperature to drop dramatically. Overall, females are relatively "inattentive" parents, incubating their eggs for only about 27-45 percent of the available daylight hours; this is significantly less than in other perching birds, who generally spend 60-80 percent. Because the eggs are laid and incubated during the coldest months of winter, they are consequently exposed to dangerously low (sometimes even sub-freezing) temperatures that generally slow embryonic development. After hatching, nestlings occasionally die from overexposure when their mother has been away from the nest for too long.
Sources
(* asterisked references discuss homosexuality/transgender)
Kenyon, R. F. (1972) "Polygyny Among Superb Lyrebirds in Sherbrooke Forest Park, Kallista, Victoria." Emu 72:70-76.
Lill, A. (1986) "Time-Energy Budgets During Reproduction and the Evolution of Single Parenting in the Superb Lyrebird." Australian Journal of Zoology 34:351-71.
* --- (1979a) "An Assessment of Male Parental Investment and Pair Bonding in the Polygamous Superb Lyrebird." Auk 96:489-98. {620}
--- (1979b) "Nest Inattentiveness and Its Influence on Development of the Young in the Superb Lyrebird." Condor 81:225-31.
Reilly, P. (1988) The Lyrebird: A Natural History. Kensington, Australia: New South Wales University Press.
* Smith, L. H. (1996-97) Personal communication.
* --- (1988) The Life of the Lyrebird. Richmond, Australia: William Heinemann.
(1982) "Molting Sequences in the Development of the Tail Plumage of the Superb Lyrebird, Menura novae-hollandiae." Australian Wildlife Research 9:311-30.
* --- (1968) The Lyrebird. Melbourne: Lansdowne Press.
Watson, I. M. (1965) "Mating of the Superb Lyrebird, Menura novae-hollandiae." Emu 65:129-32.
<<< ||||| >>>