In its embryonic origin the brain has three main areas: the forebrain, the midbrain, and the hindbrain. In the evolution of the vertebrates, the forebrain is the portion which has become most highly evolved, reaching its acme of complexity in the primates and some other higher mammals and, of course, particularly in the human animal. The chief contributions which this developed forebrain makes to human sexual behavior are: (1) an increase in the capacity to be psychologically stimulated by a diversity of erotic situations; (2) an increase in the possibilities of conditioning; and (3) an increase in the capacity to develop inhibitions which interfere with the spinal and autonomic controls of sexual behavior. This curious mixture is what many persons identify as “an intelligent control of the sexual instincts.”
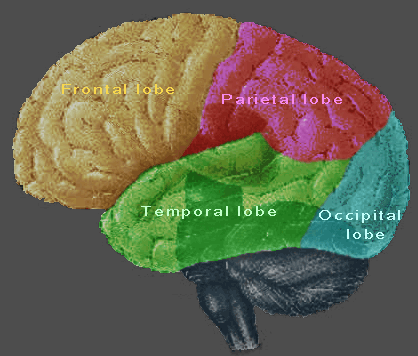
Figure 2210. Three areas and four lobes of the brain
The cerebrum of the mammal is derived from the embryonic forebrain. Sexual and emotional functions have been ascribed to certain areas of the cerebrum, including the frontal lobes, the occipital lobes, the parietal lobes, and the temporal lobes. These four areas constitute the great bulk of the human brain.
Frontal Lobes Although the frontal lobes occupy a considerable space
in the human brain, their exact function has constituted a mystery. Accidental
damage to the frontal lobes and surgical operations on the frontal lobes have
variously been reported as having no effect, or a variety of diverse and often
contradictory effects. This has been true, for instance, of the reports on the
effects of frontal lobe operations on the sexual function. There were early
reports to the effect that frontal lobe operations considerably reduced the "sex
drive” of the individual, and optimistic clinical claims that the patterns of
sexual behavior might be modified by such operations. The possibilities of such
operations have, once again, been seized upon by those who are interested in
controlling persons whose sexual activities they consider socially undesirable,
and some clinical reports have encouraged the idea that homosexual could be
changed into heterosexual patterns of behavior, that exhibitionists would lose
their compulsions to exhibit, that highly responsive, persons might become mild
and relatively unresponsive. On the other hand, there have been reports of
persons whose anxieties and inhibitions were supposed to be released and their
sexual responses increased as a result of frontal lobe operations.
For a report of reduced sexual response following pre-frontal lobotomy, see, for example: Banay and Davidoff 1942. For reports of increased sexual response following pre-frontal lobotomy, see: Hemphill 1944. McKenzie and Procter 1946. Kolb 1949 (an excellent discussion). The Columbia Greystone Project is reported in: F. A. Mettler 1949 (Selective partial ablation of the frontal cortex. Hoeber publ.) and Mettler 1952 (Psychosurgical problems. Blakiston publ.).
We have had the opportunity to make a long-range study of 95 patients who had
been subjects for frontal lobe operations. From these patients we secured
histories before operation, obtaining a record of their sexual activities for
some time prior to the operation. We similarly obtained records from these same
patients some time (a median of 3.7 years) after operation. In summary, it may be pointed out that among the females in the sample, the intensities of sexual arousal, the number of items that brought sexual arousal, the frequencies of sexual activities of particular sorts, and the frequencies of total sexual outlet did not show any significant change between a period antedating the institutionalization, and the period when the histories were retaken some years after the operation. The median lapse of time between the pre-institutional and post-institutional histories was 7.0 years.
On the other hand, among the males who had the frontal lobe operations, the frequencies of response to various stimuli and the frequencies of overt sexual activity had dropped in the course of the seven years; but the decline in responsiveness and in frequencies was not significantly different from the decline that may occur in seven years in that portion of the male population that has not had frontal lobe operations. In other words, the decline in male activity after such an operation appears to be the product of aging, rather than a direct effect of the operation. There is no decline in female responsiveness or activity because the female, unlike the male, does not show an aging effect at the ages which were involved in the experimental sample.
The critical review of the previously reported effects of frontal lobe operations, which is included in our detailed report on this material, further substantiates our conclusion that there is no demonstrated relation between the function of the frontal lobes of the human brain and any of the investigated aspects of sexual behavior. There are, inevitably, considerable shock effects from any brain operation as serious as a frontal lobe operation. Immediately following such an operation, a subject’s responses and behavior may be seriously modified. Many of the reported effects of frontal lobe operations which are to be found in the clinical literature are in actuality such immediate effects. But in most of the experimental subjects with whom we had the opportunity of working, most of these effects had disappeared within a matter of six months or so; and psychologic tests, psychiatric examinations, and a variety of other tests showed little or no permanent change in the operated sample. Some of the research group working on the project felt that there had been a lessening of emotional tensions in a statistically significant portion of the group, but this did not seem certain to some of the others working on the project, and seems not to have been reflected in any of the data which we have on the sexual behavior of the sample. The samples, however, in both our own and in most of the other studies have been small. Most of the investigators have not secured anything like precise data on the sexual behavior of the patients.
Occipital, Parietal, and Temporal Lobes
Neither is there any clearly demonstrated relation between the functions of the
occipital, parietal, or temporal lobes of the mammalian brain, and any aspect of
sexual behavior. There is one body of work that reports that operations on the
temporal lobes have a depressing effect upon emotional responses in general, but
a stimulating effect on sexual responses; but no confirmation of these results
by other investigators has yet been published.
The relative unimportance, sexually, of the parietal lobes is noted in Beach 1950:263. Lashley reports (verbal communic.) the same finding. Occipital lobe ablations in the cat produce no sexual effect other than that ascribable to blindness, according to Beach 1944:129; 1950:266. Dr. C. C. Turbes (verbal communic.) reports occipital ablations without sexual effects in dogs and monkeys. However, Klüver and Bucy 1939 describe increased sexual activity in male monkeys with bilateral temporal lobectomies; we understand that similar results were obtained with monkeys in research done under Dr. Berry Campbell at the University of Minnesota. On the other hand, Beach 1950:263-264 reports no sexual changes following bilateral temporal ablations in the cat, and Poirier 1952:234 likewise failed to find such sexual effects.
Cerebrum
On the other hand, the cerebrum as a whole seems to be significant in the sexual behavior of the mammal. Memory, various aspects of learning, various aspects of motor control, and other behavioral functions are considerably disturbed when there is accidental damage to, or operation on various parts of the cerebrum, and these may considerably affect the animal’s sexual function.
Within the last thirty years some dozen different investigators, working with a
total of six different species of mammals (rats, cats, dogs, monkeys, rabbits,
and guinea pigs), have performed operations on the cortex (the outer layers) of
the cerebrum and noted the effects of such operations on the sexual performances
of these animals.
For experiments on the relation of the cerebrum to sexual response, see: Bard 1934, 1936, 1939, 1940, 1942 (cat, dog). Brooks 1937 (rabbit). Rioch 1938 (cat). Klüver and Bucy 1938, 1939 (monkey). Davis 1939 (rat). Maes 1939 (cat). Dempsey and Rioch 1939 (guinea pig). Stone in Allen and Doisy 1939 (rabbit, rat). Beach 1940, 1942a, 1942b, 1943, 1944 (rat, cat). Langworthy 1944 (cat).
Any sort of damage to the cortex may seriously affect an animal’s motor coordination, and thus affect its physical capacity to perform effectively as a sexual partner. In addition, the cortical damage may reduce the animal’s capacity to react to psychosexual stimuli. The degree of interference is more or less directly proportional to the extent of the damage to the cortex. There are differences in the seriousness of the effects on different species of mammals.
That the extent of any interference with sexual response is correlated with the extent of cortical lesion is carefully described for the rat by: Beach 1940: 204-205, who reports diminished copulatory activity in male rats with lesions of over 20 per cent of the cortex, and a complete loss of copulatory activity when the lesion exceeds 60 per cent. But Brooks 1937:549-550 reports that male rabbits can copulate eÎFectively with all of the cortex removed, providing the olfactory bulbs are spared.
In general, cortical operations reduce the animal’s capacity to recognize (be stimulated by) sexual objects, and very much reduce its aggressiveness in approaching a sexual partner.
Localized cortical damage has been reported in a few instances to intensify sexual response, as in: Klüver and Bucy 1939. Langworthy 1944. The first authors stress the “psychic blindness” (inability to visually recognize objects) of the operated animals, but this does not harmonize with the reported “hypersexuality” which, it may be noted, seems to have been chiefly autoerotic. Likewise the frequent and protracted copulation suggests not “hypersexuality,” but an inability to achieve orgasm. The seemingly intensified responses reported by Langworthy were complicated by motor defects which interfered with effective copulation.
Since sexual relationships so largely depend on the aggressiveness of the male,
and only to a lesser extent on the aggressiveness of the female, damage to the
cortex of the male more seriously interferes with his effectiveness as a sexual
partner. Although the female with cortical damage may similarly have her
aggressiveness reduced, so that she no longer attempts to mount other females or
males, it is still possible for an intact male to mount her (if she is in
estrus) in effective copulation.
This is well demonstrated by the work of Beach. See particularly: Beach and Rasquin 1942. Beach 1943. Ford and Beach 1951:240-241.
There are a few cases of human males with cerebral damage (the exact nature of which is usually not determined) who similarly have had their responses to psychologic stimuli materially reduced by the injury. They are not aroused by any memory of previous sexual experience, and they find it difficult or impossible to explain why the previous experience was stimulating or satisfying, although they may retain some intellectual realization that the experience was formerly pleasurable. They are not aroused by discussions of sexual activities, by seeing possible sexual partners, or by seeing other sexual objects, and they show no interest in any renewal of sexual experience. However, if such a male still has his sacro-lumbar centers intact, he may still be capable of responding to direct, tactile stimulation, and he may still come to erection, copulate, and reach orgasm. There is one case of a human female who similarly had her psychologic responsiveness reduced by a cerebral injury. Unfortunately few persons with cerebral damage have had their sex histories reported.
Stier 1938 describes the sexual effects of brain injury in some 33 human males, and reports that deleterious effects were more pronounced in older males. Goldstein and Steinfeld 1942 give a detailed discussion of a single case. Both papers note a marked reduction in response to psychologic stimuli. The female case was originally reported by Symonds and is cited in other literature, e.g., Beach 1942a :216.
Although the data on the relation of the cortex to sexual behavior are limited,
they do show that this is the part of the nervous system through which
psychosexual stimuli are mediated. Since there are considerable differences in
the effectiveness of such psychologic stimuli between females and males, we may
believe that this, the most striking disparity which exists between the
sexuality of the human female and male, must depend on cerebral differences
between the sexes. What the nature of such cerebral differences may be, we do
not know. There have been one or two studies which report differences in the
biochemistry of the cerebral cortex in female and male animals. The studies are
important and highly suggestive, but further investigation is needed before we
are warranted in making any generalization.
For studies of chemical differences that appear to distinguish female from male brains, in both human and infra-human species, see: Weil 1943. Weil and Liebert 1943. Weil 1944.
Since there are differences in the capacities of females and males to be conditioned by their sexual experience, we might expect similar differences in the capacities of females and males to be conditioned by other, non-sexual types of experience. On this point, however, we do not yet have information.
Hypothalamus While we may explain the similarities of female and male responses to tactile stimulation on the basis of the similarities of spinal and autonomic mechanisms, and while we are inclined to believe that differences between female and male responses to psychologic stimuli may depend on cerebral differences, we still have one of the most significant aspects of sexual response to explain. This is the fact that sexual responses constitute a syndrome of elements most of which are found in other emotional responses, although the combination in which they appear during sexual activity is not duplicated in any other type of behavior. It is still difficult to understand why touching an animal at one time should bring responses which we recognize as sexual, while touching it on some other occasion may make the animal angry or afraid.
It is inevitable that one should assume that such a cluster of responses must depend upon some mediating mechanism, a master switchboard which controls all of the individual elements but brings them together as a unit during sexual response. Perhaps it is not reasonable or necessary to believe that there should be such a mediating mechanism, but the possibility is sufficient to warrant continued search for a central control of the sexual syndrome.
Such '‘sex centers” in the cerebrum have been postulated by various students, including: Loewenfeld 1908:597-598. Rohleder 1923:4, 19. Von Bechterew acc. Beach 1940:194. A summary is in Stone 1923b. However, recent and thorough research has discovered no such centers, and Penfield and Rasmussen 1950:26 stress that they were never able to elicit erotic sensations by stimulating the exposed human brain.
The concept of a master switchboard is encouraged by experimental work which
shows that damage to the hypothalamus, a small structure in the brain which lies
below the cerebrum, may considerably modify the animal’s capacity to be aroused
in anger, in fear, and in still other emotional responses. This it seems to do
through some control which it exerts on the autonomic nervous system.
Nevertheless, those who have done the most extensive research on this particular
portion of the brain conclude that there is no evidence—either for or
against—that the hypothalamus in any way controls sexual responses. The
statement was originally made some twenty years ago, but we are advised that it
still represents our present state of knowledge or lack of knowledge on this
matter.
The role of the hypothalamus in mediating autonomic elements and in the expression of emotion is noted by: Fulton 1949:237, 243-245. The sexual role of the hypothalamus is emphasized by some, as in: Brookhart and Dey 1941. Dey, Leninger, and Ranson 1942. Ford and Beach 1951:240. But Clark 1942 and Bard 1940 report inconclusive results and feel that the evidence is still insufficient for final judgment. Bard 1940:574, 576, states “There are not yet available sufficient experimental facts to warrant any general statement about the relation of the hypothalamus to the excitation and execution of estrual behavior. . . . Further work must be done before any precise statement can be made concerning the relation of the hypothalamus to the central management of sexual behavior.” This author recently (verbal communic.) considers the above statements still valid.
We have made considerable progress in understanding the anatomy and the gross physiology of sexual response. After three decades of research done by a score of students of human and infra-human sexual behavior, we are able to identify some of the internal mechanisms which account for the similarities between female and male sexual responses, and have located the portion of the brain which seems responsible for the differences which we have found in the capacities of females and males to respond to psychologic stimuli. With this much of the story pieced together, it should now be possible for the observer of gross behavior, the anatomist, the neurophysiologist, and the student experimenting with mammalian sexual behavior to recognize the areas in which we most need additional research.