|
|
|
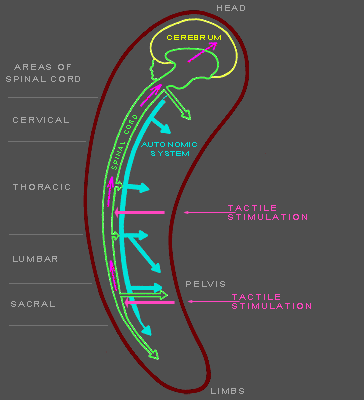
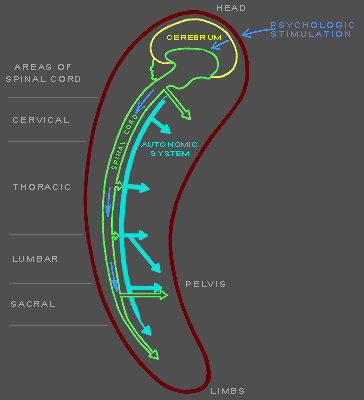
Figures 141f-142f. Diagrams of neural mechanisms in mammalian |
|
|
|
|
|
Figures 141f-142f. Diagrams of neural mechanisms in mammalian |
|
|
| Table 176f. Physiologic Elements in the Sexual Syndrome, Anger, Fear, Epilepsy, and Pain |
| Physiologic element | Neural mech. | Sexual resp. | Anger | Fear |
Epi- lepsy | Pain |
| Increase in pulse rate | symp. | v | v | v | may | v |
| Increase in blood pressure | symp.? | v | v | v | may | v |
| Vasodilation | para. | v | may | no | v | |
| Increased periph. circ. of blood | para. | v | may | no | may | |
| Tumescence | para. | v | rare | rare | rare | rare |
| Reduced rate of bleeding | symp. | v | v | v | v | |
| Hyperventilation | symp. | v | v | v | v | |
| Breathing irregularity | v | v | v | may | v | |
| Anoxia | ? | v | ||||
| Diminished sensory perception | v | v | v | v | v | |
| Increase in genital secretions | v | no | no | |||
| Increase in salivary secretion | para. | v | v | no | v | no |
| Increase in lacrimal secretion | v | v | ||||
| Increase in perspiration | para.+symp. | occas. | v | v | ||
| Adrenaline secretion | symp. | v ? | v | v | v | |
| Increase in muscular tensions | spinal | v | v | v | v | v |
| symp. | ||||||
| Increase in muscular capacity | v | v | v | |||
| Involuntary muscular activity | spinal | v | v | v | v | v |
| Rhythmic muscular movements | spinal | v | no | no | may | no |
| Gastro-intest. activ. inhibited | symp. | v ? | v | v | v | |
| Hair raised | symp. | may | v | v | may | |
| Eye pupil dilated | symp. | v | v | v | v | v |
| Ejaculation | spinal | v | rare | rare | may | |
| symp. | ||||||
| Involuntary vocalization | v | v | v | v | v | |
|
v = physiologic element is present. No = element is absent. May = element sometimes but not always present. | ||||||