Among the internal factors which may affect the way in which the animal body functions, the best understood are the hormones. These are chemical substances which are produced chiefly in endocrine organs, from which they are ultimately carried by the blood stream to every part of the vertebrate body. Because of their accessibility to all parts of the body, hormones may have more effect on bodily functions than any other mechanism except the
nervous system.
Among all of the hormones that are now known to exist in the vertebrate body, the so-called sex hormones were practically the first to be discovered.
It is probable that most races of men, including even the most primitive, have recognized that the testes served two roles—one in connection with reproduction, and one in connection with the growth and function of the body as a whole. Long before it was known that the ovaries and testes produce specific reproductive cells, the eggs and the sperm, it was known that some substance produced by the male had to be transferred to the female tract before she could reproduce; and the inability of a male to contribute to such reproduction after his testes had been removed provided early evidence that they were the source of an essential part of this fertilizing substance.
The Trobriand Islanders and certain Australian tribes are said not to be aware of the relationship between coitus and pregnancy, but even they consider that coitus paves the way for subsequent supernatural impregnation. The Trobrianders castrate boars “to improve their condition” and note that such boars cease copulation. This renders suspect Malinowski’s statement that the testes are considered to be only ornamental appendages. See: Malinowski 1929:168, 180-181, 190-191. Ashley Montagu 1937:199-202.
Although there was, of course, no understanding that a chemical mechanism was involved, both primitive and ancient peoples also recognized that the early physical development of the male animal and its capacity to engage in sexual activity also depended upon its possession of testes. Human male castrates as well as castrates among farm animals were known to the earliest peoples in all parts of the world, and consequently the effects of castration were well understood at an early date. Because the ovaries are within the body cavity, female castrations are more difficult to perform, and were rarely done before the days of modern medicine. Consequently the significance of the ovaries in regulating body functions was not understood at as early a date.
The understanding and practice of castration may have begun as early as 7000 B.C. in the early Neolithic when animals were first domesticated. See: Steinach 1940:3, 25. Turner 1948:324. Many primitives who were at a Neolithic cultural level when first visited by Europeans practiced castration. See, for instance: Malinowski 1929:191 (Trobriand Islanders). Evans-Pritchard 1940:33 (Nuer of Africa). It is interesting to note that in cases of self-castration, as on the islands of Ponape and Tonga (Westermarck 1922(1):561) and among die Hottentot of Africa (Bryk 1934:123), the males were careful to remove only one of the testes. In addition, there are references to castration in some of the oldest myths of Europe and Egypt. See: Möbius 1903:12. Pritchard 1950:181 (Middle Assyrian laws, 15th century b.c., tablet A, laws 15 and 20, provide castration as a punishment).
The ancients knew that the effects of castration depended upon the age at which the human or other male was castrated. They knew that when the testes of the human male were removed before the onset of adolescence, the effects were more marked than they were when the castration was performed on a male who was a fully grown adult. 3 The basic biology of these matters did not find an explanation, however, until the middle of the nineteenth century.
Aristotle (384-322 b.c.), in the Historia Animalium, Bk. IX:631b-632b, has a lengthy discussion of the effects of castration in man and in other male animals, differentiates between pre-adolescent and adult castration, and refers to the removal of the ovaries of sows to lessen their sexual responsiveness.
In 1835, Graves recognized the relation between thyroid pathologies and the physiologic disturbances which accompany the disease which now bears his name. In 1849, Berthold, studying castrations and testicular implants in fowl, concluded that the testes secreted one or more blood-borne substances which were responsible for the modifications which his experimental castrations had produced. Within the next decade, Addison had noted the deterioration of the adrenal cortex in victims of the disease which bears his name. By the mid-seventies. Gull had identified the role of the thyroid in certain pathologic conditions; by 1887 Minkowski associated acromegaly with pituitary hyperfunction, and two years later (in 1889) von Mering and Minkowski had removed the pancreas and experimentally produced diabetes. The work of Brown-Séquard in 1889 revived interest in the utilization of testicular extracts in clinical practice, and since then there has been a tremendous development of experimental work on the significance of both testicular and ovarian hormones.
For brief histories of endocrinology, see: Corner 1942:228-233. C. D. Turner 1948:5-9.
The list of endocrine glands which are now recognized in the vertebrate body include the ovaries and testes (or the
gonads, as the two sets of organs are generically called), the pituitary gland which is located at the base of the brain, the thyroid and parathyroid glands which are located in the throat, and the adrenal glands which are located at the top of the kidneys near the small of the back. The thymus is a gland which reaches its maximum development in the early life of the animal but degenerates considerably after that.
The removal of the thymus and/or injection of thymus extracts are usually without effect; but the development of the gonads at the onset of adolescence is associated with an atrophy of the thymus, and castrates retain the large thymus of childhood. See: Hoffman 1944:275. C. D. Turner 1948:21-22. Selye 1949:679, 683.
The pineal gland, in the brain, may be an endocrine organ.
The functions of the pineal gland are poorly known and the data are conflicting, but see: Hoffman 1944:282-284. C. D. Turner 1948:19-21. Selye 1949:595.
The liver and the pancreas, in addition to secreting substances which directly affect digestion, also produce important hormones which influence the development and the maintenance of activities of other organs of the body.
It is not fully demonstrated that the liver secretes a hormone, but secretions of the liver inactivate gonadal and probably other hormones. See: C. D. Turner 1948:25. Williams 1950:7.d
The testes are the chief source of the several hormones known as androgens (the so-called male hormones) in the body of the male. One of the best known androgens is testosterone. The ovaries are the chief source of the so-called female hormones in the female. The most prominent of the female hormones are the estrogens and progesterone. As a group, hormones from the ovaries and testes may be referred to as gonadal hormones.
The estrogenic hormones found in human tissues and fluids are estradiol, estrone, and estriol. Estradiol is thought to be the true hormone and the others to be products derived from it, according to: Smith in Williams 1950:351. Talbot et al. 1952:296.
In recent years, the hormones produced by most of the endocrine glands have been isolated and identified as specific chemical substances. Many of them are closely related compounds of carbon, hydrogen, and oxygen. For instance, the androgens and estrogens, the 17-ketosteroids which are produced by the adrenal glands and by some other structures in the body, and the steroids which are chemicals characteristically found in all animal tissues, are all closely related chemical compounds, even though each may have a different and unique effect on the physiology of the body. Some of the other hormones, such as those produced by the thyroid and the pituitary glands, are totally different in their chemical composition.
Nature of the Hormones A general knowledge of the hormones has become widespread in the population as a whole, but in regard to certain critical matters this knowledge is quite incorrect. Journalistic accounts of scientific research, over-enthusiastic advertising by some of the drug companies, over-optimistic reports from clinicians who have found a lucrative business in the administration of sex hormones, and some of the discussions among state legislators and public administrators who hope that hormone injections will provide one-package cure-alls for various social ills, have led the public to believe that endocrine organs are the glands of personality, and that there is such an exact knowledge of the way in which they control human behavior that properly qualified technicians should, at least in the near future, be able to control any and all aspects of human sexual behavior. It is, therefore, important that the general reader understand the nature of the hormones, and understand some of the difficulties that are involved in the accumulation and interpretation of data in the field of endocrinology.
Hormones are products of the physiologic processes that go on in certain of the gland cells that are to be found in the plant or animal body. Any cell which secretes a liquid content which becomes a significant part of the total volume of the cell, or which secretes materials which work their way out of the cell through a permeable cell wall or through some rupture of the wall, may be identified as a gland cell. Many of the cells of the body, and particularly those that line various body cavities, may be considered gland cells even though they are not part of a specific organ which is identifiable as a gland. Consequently, it is not always possible to identify all of the sources of the hormones in an animal’s body, even including the androgens and estrogens and the 17-ketosteroids, for part of the hormone may come from cells or groups of cells which lie outside of the specific organs which are known to be the chief sources of these hormones. For instance, the removal of the ovaries or testes (as in a complete castration) may not eliminate all the sources of the sex hormones, and this is one reason that it is difficult to interpret some of the experimental data.
For instance, after removal of the testes, the cortex of the adrenal gland enlarges and secretes an additional amount of androgenic (male) hormone in mice and guinea pigs. See: Hartman and Brownell 1949:331. Selye 1949:130-131.
In more complex glands, the secreting cells may pour their products into internal cavities from which ducts may carry them away. This is true, for instance, of the salivary glands. On the other hand, the glandular structures which give rise to the best known of the hormones do not have either internal cavities or ducts. Their secretions are picked up by the blood vessels which enter or surround the glands, and are thus carried away by the blood stream to other parts of the body. The structures are therefore known as ductless glands, or glands of internal secretion, or endocrine (meaning
internally secreting) organs. There are, however, hormones produced by gland cells in other types of structures, such as the placenta and the duodenal mucosa; and there is some reason for believing that most of the organs in the mammalian body may produce, in actuality, substances which, when circulated through the blood stream, may influence the activities of at least some of the other organs.
The placenta secretes estrogen, progesterone, and a gonadotropin. The duodenal mucosa secretes several hormones concerned with digestion. See: C. D. Turner 1948:13-16.
The hormones produced by any endocrine gland may affect other endocrine glands as well as organs which are not glandular. For instance, the secretions of the testes and ovaries have a direct effect on the anterior lobe of the pituitary and on the adrenal glands, and each of these has a direct effect on the testes and ovaries. Consequently an increase or decrease in the secretory capacity of any one of these glands may be reflected in the activities of the other glands. Some of the other endocrine organs, such as the thyroid, may similarly affect the secretory capacities of the ovaries and testes, and of the pituitary and adrenal glands.
For the interrelationships between the secretions of the anterior lobe of the pituitary, the ovaries, testes, and adrenal cortex, see: Heller and Nelson 1948: 229-243. C. D. Turner 1948:241-244, 462-468. Hartman and Brownell 1949: ch. 26. Selye 1949:22-23. Burrows 1949:329-343. Williams 1950:21-32. Talbot et al. 1952:385-392.
Although the effectiveness of any hormone is usually proportionate to the amount which is available, there is usually a point of optimum effectiveness, and an increase in the amount of hormone beyond that point may have negative effects which, in certain respects, may be as extreme as those obtained when there is an under-supply or complete removal of the source of supply of the hormone.
For instance, both a thyroid deficiency and an excess of thyroid retard sexual maturation. See: Lisser 1942:29-32. Hoffman 1944:239-240. Williams 1950: 122, 175. Talbot et al. 1952:21, 31. See footnotes 37, 66.
Usually the amount of hormone produced in an endocrine organ such as a testis or ovary is very small, and the amount that is to be found in the blood or the urine or at any other point in the body is so exceedingly minute that its recovery and chemical identification may be very difficult. Consequently, most of the reports of female and male hormone levels do not rest upon physical or chemical measurements, but upon such indirect evidence as can be obtained by injecting urine or blood extracts into experimental animals, and upon measurements of the changes which are thus effected in the growth or degeneration of some structure (like a roosters comb) in the experimental animal. Difficulties in measurement have been the source of considerable error in much of the reported work, including the studies which have attempted to analyze the relationships of the sex hormones and sexual behavior.
As an example of the difficulties encountered in analyses: testicular androgen is metabolized and excreted partly in the form of ketosteroids in the urine, and hence the injection of an androgen such as testosterone propionate may be followed by an increase in urinary ketosteroids. But another androgen, methyl testosterone, may have no effect on the levels of the urinary ketosteroids, or even decrease them. See: Selye 1949:628. Howard and Scott in Williams 1950:337.
Moreover, when the amount of hormone in an animal’s body is determined by measuring the hormone in its urine, it is questionable what relation the amount of excreted material may have to the amount of the hormone that the body is actually utilizing. The hormone in the urine may merely represent that portion which the body has been unable to utilize. Recent endocrinologic research indicates that this latter may be the correct interpretation, especially when an animal is receiving an over-supply of hormones. In an effort to allow for this, hormonal measures are often made on materials recovered from the blood; but it is not clear that this eliminates the difficulty, for it is still not certain how much of the hormone carried in the blood stream may ultimately be utilized by an animal. Consequently reports on hormonal levels in females and males, or in heterosexual and homosexual males, are acceptable only when allowances are made for possible errors in interpreting the measurements, when a sufficient allowance is made for variation in the same individual on different occasions, when the series of reported cases is of some size, and when the report is based on a statistically adequate experimental group which can be compared with an adequate control group.
Development of Physical Characters in Young Mammals
The most certain effects of the gonadal hormones are their effects on the development of physical characters in the young animal. This includes the development in size and function of many parts of the body, including some of the characters (the secondary sexual characters) which most clearly differentiate adult females from males. Many of these characters do not fully differentiate until the onset of adolescence. The development of these secondary sexual characters, as well as the development of the adult anatomy as a whole, depends upon the animal’s possession of intact gonads. This is true of the human female and male, and of the females and males of the lower species of mammals.
If the gonads fail to develop normally—as not infrequently happens when testes of the human male, for instance, are retained in the body cavity and fail to descend into the scrotum—or if the gonads of the pre-adolescent human female or male become diseased, or if they are removed by castration any time before the onset of adolescence, the normal development of adult characters is usually slowed up or completely stopped. And when a castrated animal does reach its full size, its body proportions are not typical of those usually found in the normal adult, i.e., the animal becomes a typical capon, a gelding, or a eunuch in form and structure.
The failure of pre-adolescent castrates, either surgical or functional, to develop the secondary sexual characters typical of the adult, is described in such standard works as: Laughlin 1922:435. Lipschütz 1924:6 ff. Pratt in Allen et al. 1939:1267-1268, 1282. Hamilton 1941:1904. Greenblatt 1947:182, 254-257. Mazer and Israel 1951:231. Ford and Beach 1951:170.
In the normal human adolescent, female or male, the genitalia are among the first structures to acquire adult size. When the gonads have been damaged before adolescence, the genitalia may develop even more slowly than the rest of the body and may remain more or less infantile even into later years.
For data on the growth of the human male genitalia before the rest of the body is fully grown, see: Schonfeld and Beebe 1942:771.
In the human female and male, hair normally begins to develop in the armpits during adolescence. In normal development, pubic hair appears in both sexes, although the pubic hair of the female is ordinarily confined to a more limited triangle while it may spread over a more extensive area in the male. Ultimately, but often not until late in the twenties or thirties, the pubic hair of the male may develop upward along a midline (the
linea alba) on the abdomen. The normal male also develops hair on his face, on his chest, on his legs, and elsewhere on his body, while such hair is usually absent or scant in the female. But if the testes of the pre-adolescent male are damaged or eliminated, the hair in these several parts of the body may fail to appear at the usual age. If it does subsequently develop in the castrated male, it may appear in a pattern which is in many respects more typical of the very young adolescent. The face and chest and other parts of the body of such a male may remain more or less hairless.
Absence of pubic hair and abnormal hair distribution in males with gonadal insufficiencies are well illustrated in Selye 1949:651-660.
While the early castration of a female does not have as marked an effect on the development of her body hair, it may contribute to the appearance of facial hair and other hair developments which are not typical of her sex.
A diminution of axillary and pubic hair, and sometimes hirsutism in females with congenitally absent or undeveloped ovaries are noted in Selye 1949:399-400.
The Adam’s apple is characteristic of the adult human male. Associated with this, his voice is rougher and usually at a pitch which is lower than that characteristic of the female. The Adam’s apple is ordinarily not developed in the female. The male who is castrated before adolescence fails to develop an Adam’s apple and retains a high voice.
In the course of adolescent development the shoulders of a normal male widen more than they do in the normal female. In adult females the hips become characteristically larger and wider than they are in the male. The buttocks in an adult female usually become larger and more elongated, while the buttocks of the adult male remain smaller and are more often rounded. The male who is castrated before adolescence retains body proportions which are closer to those of the juvenile.
The breast of the female normally enlarges in size, and the colored, corrugated (areolar) area surrounding the nipple becomes considerably expanded. The female who has diseased or damaged ovaries does not show such a normal breast development, her voice may become lower in pitch, and she may fail to acquire an adult female body form.
In general, castrations performed on young females and young males of the infra-human species of mammals affect their physical development in ways which are comparable to those just noted for human females and males.
Some of the effects of castration may be partially or largely corrected by the administration of hormones from an outside source. This is true for both females and males, and for both the human and other mammalian species. But injections of hormones have their maximum effect if they are made at an early age. They cannot fully correct the damage done by a castration if they are not administered until some time after adolescence has begun, but they may still have some value even when the therapy is not started until the individual is essentially adult. But the corrective effects of hormonal administrations to young castrates can be maintained only if the treatment is continued throughout the growth period. Otherwise the individual may lapse into its castrated state. If hormonal treatments are continued until the individual has become completely adult—which in the human species means into the middle twenties or sometimes later—then the continued administration of hormones is not so necessary.
In the abundant literature on the correction of a gonadal insufficiency through the administration of hormones, see, for instance: Kenyon 1938:121-134. Vest and Howard 1938:177-182. McCullagh 1939. Hamilton and Hubert 1940:372. Escamilla and Lisser 1941. Biskind et al. 1941. Kearns 1941. Moore 1942:39. Heller, Nelson, and Roth 1943. Hurxthal 1943. Heller and Maddock 1947:414-418. Beach 1948:38-41, 45. Selye 1949:645-666. Howard and Scott in Williams 1950:328-333. H. H. Turner 1950:32-48.
Maintenance of Physical Characters in Adults
Damage to the gonads, or a complete castration of a human female or male after physical maturity has been acquired, prevents reproduction, but there are usually minimum effects on other physical characters. Some individuals (particularly some females) who have been castrated as adults may go for years and may even reach old age before they show any marked physical changes. Some females and males, on the contrary, may show more physical deterioration in the course of time. Clinicians commonly report characteristic aging effects on the genitalia of females who have had their ovaries removed. It is generally believed that the deteriorations of old age come on sooner, although the specific data are inadequate on this point.
Menopausal-like involutional changes in the genitalia of castrated adult females, and their control by estrogen therapy, are noted, for example, in: Hoffman 1944:33-35. The lack of recessive changes in some hypogonadal males is discussed in: Heller and Maddock 1947:395.
Castrations may, however, have marked effects on the physiologic well-being of an adult. Since gonadal secretions affect the levels of secretion of the pituitary, adrenal, and thyroid glands, all of which are important in the regulation of the general physiology of an animal, it is inevitable that castrations, even of adults, should have some effect; but this effect is usually minor among human females, and it is not clear that most human males have their physiologic well-being particularly modified by castration if the operation is performed after complete physical maturity has been acquired.
The general physiologic effects of castration are discussed, for instance, in: Möbius 1903:28 ff. Laughlin 1922:435-436. Lipschütz 1924:12-14. Wolf 1934:257-268, 279. Lange 1934. Greenblatt 1947:254-257. Heller and Mad-dock 1947:393 ff. Ford and Beach 1951:221-225, 229-232.
There seem to be more marked effects on the physical characters and on the
physiologic well-being of males of lower mammalian species which are castrated
as adults.
In animals, adult male castrates show a genital atrophy, an accumulation of fat, and a decrease in metabolism, sexual drive, and aggressiveness. See: Tandler and Grosz 1913:25-41. Rice and Andrews 1951:115-116.
The effects of castration on an adult animal, such as they are, may be more or
less completely corrected by the administration of a sufficient supply of
gonadal hormones. This has been demonstrated for laboratory animals, and
hormones are often administered in clinical practice to middle-aged and older
women who have had their ovaries removed. Usually testosterone is given to a
male who is castrated as an adult, and estrogens to a female, but sometimes both
hormones are given to individuals of both sexes. It is significant that the
corrective administrations of hormones do not need to be kept up indefinitely in
an adult, at least in an adult female. In some way the adult human body can
adjust in a matter of months to a lack of gonadal hormones, and then it appears
to be capable of more or less normal function, even though an important link in
the endocrine chain has been eliminated. The capacity of the adult male body to
adjust may not be as complete as that of the female. Some adult male castrates
appear to adjust to a lack of male hormones for long periods of years; but
others show some physical degeneration within a shorter period of years. Until
there are further studies of long-time adult male castrates, we are uncertain
how to interpret these contradictions in the reported data.
Levels of Gonadal Hormones and Sexual Behavior
Much of the confusion concerning the function of the hormones which originate in the ovaries and the testes is a consequence of the unwarranted opinion that anything associated with reproduction must, ipso facto, be associated with an animals sexual behavior and, contrariwise, that all sexual behavior is designed to serve a reproductive function. Since the glands which produce eggs and sperm also produce hormones, scientists and philosophers alike have considered it logical to believe that these must be the hormones which control sexual behavior. Reasoning thus, men throughout history have castrated criminals as punishment for sexual activity which their gonads were supposed to have inspired, and with the intention of controlling their further sexual activity.
For example, in the Middle Assyrian laws, which may date back to the 15th century B.C., tablet A, laws 15 and 20, provide castration as a punishment for certain sexual offenses. See: Pritchard 1950:181.
In recent years, courts and state legislatures are again considering gonadal operations as a means of controlling sex offenders.
There are statutes in some ten states providing for involuntary asexualization, or sterilization as a eugenic or therapeutic measure for criminal conduct. In several of these states (e.g., Kansas and Oregon) the practice has been to allow castration as one of the permissible operations. Since the United States Supreme Court decision in Skinner v. Oklahoma 1941:316 U.S. 535, all operations have been “voluntary.” In order to circumvent constitutional objections, the California Legislature at its Third Extraordinary Session in 1951 passed a bill, A.B.2367, providing for mandatory life imprisonment for persons convicted of certain types of criminal conduct who did not “consent” to being castrated. This bill was vetoed by Governor Warren on July 18, 1951. A milder version of the same bill, S.B.19, failed to pass in the 1952 regular session of the Legislature. We are informed that a similar proposal was introduced in the 1953 session of the Oregon Legislature. To our knowledge castration proposals were summarily rejected by the commissions on sex offenders in New Jersey and Illinois. See also, Michigan, Governor’s Study Commission 1951:4, 6.
There has been some experimentation with hormone injections in an attempt to achieve that end. Castrations and the administration of sex hormones have been carried out under court order and under the direction of physicians and psychiatrists in various parts of the United States, in Denmark, in Holland, and still elsewhere in mental and penal institutions.
For such a use of castration or the administration of hormones, see: Hirschfeld 1928:54. Lange 1934:44, 101. Wolf 1934:16-23. Böhme 1935:10-34. Sand and Okkels 1938:374. Kopp 1938:698-704. Hawke 1950. Tappan 1951:242-246 (Denmark). Bowman 1952:70 (cites Judge Turrentine of San Diego court as stating that “behavior disorders have been well controlled by castration in about 70 men,” and that these men failed to become involved again with the law after castration. Since sex offenders are always among the lowest in their rate of recidivism, the Judge’s criterion is inadequate.). Bowman 1952:79-80 gives a report from the Swedish authorities on 166 legal castrations between 1944 and 1950. Bowman and Engle 1953:10.
On the even more amazing assumption that anatomic defects in the genitalia may explain the social misuse of those organs, some of the medical and psychiatric officers in police courts and in penal and mental institutions routinely examine the genitalia of persons committed on sex charges.
But the fact that hormones are produced in the gonads is, without further evidence, no reason for believing that they are the primary agents controlling those capacities of the nervous system on which sexual response depends. It is unfortunate, as we shall see, that these hormones were ever identified as sex hormones, and especially unfortunate that they were identified as male and female sex hormones, for the terminology inevitably prejudices any interpretation of the function of these hormones.
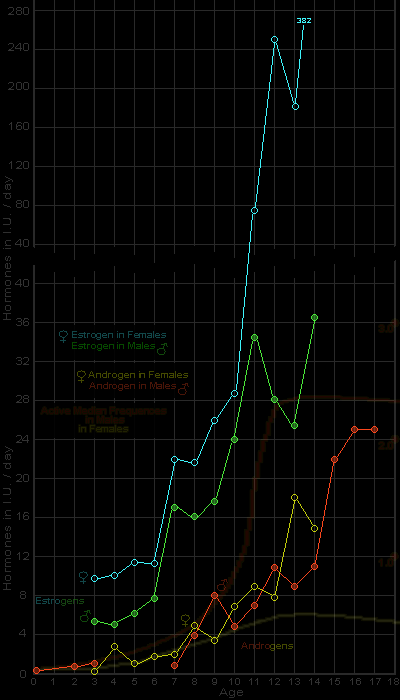
Figures 152f-153f. Androgen and estrogen levels in pre-adolescent and adolescent female and male
In comparison with active median frequencies (per week) of orgasm in total sexual outlet of males and females (Figure 143f). Averages from urinary assays reported by Dorfman 1948 and Nathanson et al. 1941
Estrogen Levels at Younger Ages
It should be borne in mind that estrogens (the female hormones), are to be found in the bodies of both females and males. The ovaries of the female are a chief source of her estrogens. The origins of the estrogens in the male are not so well established, but they seem to be produced, at least in part, by the testes.
For data on the two estrogens, estradiol and estrone, found in the testes, see: Selye 1949:54, 84. For data on estrogenic substances from the adrenal cortex, see: Hartman and Brownell 1949:105. Selye 1949:80-81. Kepler and Locke in Williams 1950:203. Thom and Forsham in Williams 1950:261.
Estrogens are reported to occur in about equal amounts in the preadolescent human female and pre-adolescent human male until they reach the age of ten (Figure 152). But at about the time of adolescence, the estrogens increase, abruptly in the female. There is only a slight increase in estrogens in the male at adolescence.
For levels of gonadal hormones in pre-adolescence and adolescence, see Nathanson et al. 1941.
In the adult female there is, in consequence, a much higher estrogen level than in the adult male. There is, of course, wide individual variation in this matter.
For androgen and estrogen levels in normal adult males and females, see: Gallagher et at. 1937:695-703. Koch 1938:228-230. Heller and Maddock 1947: 395-398. Dorfman in Pincus and Thimann 1948:496-508.
There is nothing, however, in the development of sexual responsiveness and activity, either in the female or in the male, which parallels these reported levels of estrogens in the human female or male. At the onset of adolescence there is no upsurge of sexual responsiveness and sexual activity in the female which parallels the dramatic rise in the levels of her estrogens (Figure 152f). It is the male who suddenly becomes sexually active at adolescence, but his estrogens stay near their pre-adolescent levels.
Androgen Levels at Younger Ages Androgens are also found in both females and males. The testes of the male are the chief source of his androgens; but the ovaries of the female apparently produce androgens as well as estrogens, and it is probable that the adrenal glands and still other structures in her body also produce androgens.
The production of androgenic substances by the ovary is noted in: Hoffman 1944:47. C. D. Turner 1948:337. Burrows 1949:123-124. Selye 1949:627. Parkes 1950:108. The adrenal cortex as another source of androgens is noted in: Koch 1938:218. Nathanson et al. 1941:862. C. D. Turner 1948:337. Burrows 1949:120. Selye 1949:80, 126-127. Kepler and Locke in Williams 1950:203. Parkes 1950:102. Perloff in Mazer and Israel 1951:124.
In the human species, from about age seven or eight until the middle teens, the androgen levels in the female and the male are about equal (Figure 153). Then the androgen levels begin to rise more markedly in the male, and less so in the female, and it is generally considered that older females have androgen levels that are about two-thirds as high as those of the males.
Since we found a sudden upsurge of sexual responsiveness and overt sexual activity among human males at the beginning of adolescence, there may seem to be some correlation with the androgen picture; but the upsurge of sexual responsiveness in the male is much more abrupt than the steady rise in the levels of his androgens (compare Figures 143f and
153f).
As for the female, there seems to be no correlation at all between the levels of her androgens and her slow and gradual development of sexual responsiveness and overt sexual activity (Figure 143f and
153f). Although she has nearly as much androgenic hormone as the male in her pre-adolescent and early adolescent years, her levels of sexual response and overt sexual activity at that period are much lower than the levels in the average male. The near identity of the androgen levels in the female and male at the very age at which the two sexes develop such strikingly different patterns of behavior, makes it very doubtful whether there is any simple and direct relationship between androgens and patterns of pre-adolescent and adolescent sexual behavior in either sex.
Levels of Gonadal Hormones in Older Adults Unfortunately, levels of gonadal hormones seem not to have been established for any adequate series of older human adults, either female or male. There is some reason for assuming that the levels of male hormones drop in the male at advanced ages, and if this were proved to be so it would parallel the drop in the levels of sexual response and overt activity which we have found in the male. On the other hand, there is as much reason for assuming that the levels of male hormone similarly drop in the older female, but such a drop would not correlate with the fact that the frequencies of female response and sexual activity stay on a level from the teens into the fifties or sixties (Figure 143f).
Dorfman in Pincus and Thimann 1948:502-504 differentiates the androgens from the 17-ketosteroid data and shows a decline in androgens with advancing age in both female and male. Ford and Beach 1951:227 state that it is generally agreed that testicular androgens decrease in later life.